
+91 6002993949
submission@iarconsortium.org
Open Access
ISSN (Print) : 2788-9475
ISSN (Online) : 2788-9483
Background: Histamine is a biogenic amine produced from different tissues and organs as skin, lung and digestive system. It play important role in regulation of sensation of pain, body temperature, memory, and wakefulness. Objectives: The current review aimed to highlight on the structure, metabolism and the physiological effects of histamine. Histamine is synthesized from the L‐histidine amino acid in both the neuronal cell bodies and axon terminals and stored in vesicles in axon varicosities. In the brain, histamine is also produced by mast cells. The release and synthesis of histamine are regulated by H3 receptors. The action of histamine is modulated by four types of G protein coupled receptors (H1, H2, H3, and H4). The histamine releasing and synthesizing neurons are found in tuberomamillary nucleus in hypothalamus. It is a key player in allergic reactions, circadian cycle regulation, vascular permeability, gastric and neurological functions, immune cell differentiation, epithelium proliferation, neoplastic progression and angiogenesis. Release of toxic substance from ischemic tissue as tissue enzyme, histamine, and serotine is the main cause of circulatory shock. In inflammatory responses and allergic reactions, histamine causes arterioles vasodilator and increases the porosity of capillaries to leak plasma and plasma proteins into interstitial space leads to edema. The rate of formation and secretion of hydrochloric acid by the parietal cells is directly related to the amount of histamine secreted by the enterochromaffin-like cells, that may be stimulated by hormones secreted by the enteric nervous system of the stomach wall. Histamine and slow reactive substance of anaphylaxis released in the lung tissues during allergic reactions by tissue mast cells by the action of allergic substances as pollen grains, irritant chemicals or vapors and cause bronchial constriction. Histamine intolerance results from a disequilibrium of accumulated histamine and the capacity for histamine degradation. Ingestion of histamine-rich food, alcohol, or drugs that release histamine or block diamine oxidase may provoke diarrhea, headache, congestion of the nose, asthmatoid wheezing, hypotension, arrhythmia, urticaria, pruritus, flushing, and other conditions in these patients. Conclusion: It can be concluded that the action of histamine is modulated by H1, H2, H3, and H4 receptors. It is a key player in allergic reactions, circadian cycle regulation, vascular permeability, gastric and neurological functions, immune cell differentiation, epithelium proliferation, neoplastic progression and angiogenesis. The rate of formation and secretion of hydrochloric acid by the parietal cells is directly related to the amount of histamine secreted by the enterochromaffin-like cells. The release of histamine may be provoke diarrhea, congestion of the nose, headache, asthmatoid wheezing, hypotension, arrhythmia, urticaria, pruritus, and flushing.
Histamine is a highly pleiotropic biogenic amine involved in key physiological processes including neurotransmission, immune response, nutrition, and cell growth and differentiation [1]. It is produced from different tissues and organs as skin, lung and digestive system, it also stored in granules of mast cells and basophiles in the blood in association with heparin. It found in entrochromaphin- like cells in high concentration in lymph nodes, thymus and gastric glands and in intermediate concentrations in lung, liver and varicosities of histamine releasing neurons. Histamine acts as a local mediator in gut and immune system and a neurotransmitter in nervous system to bring about many physiologic changes as, inflammation, smooth muscle contraction in uterus, lung, and intestine, vasodilatation and modulation of heart contractility and rat, hydrochloric acid secretion in stomach, production of cytokines and chemotaxis [2].
Histamine also has many functions in nervous system in regulation of body temperature, sensation of pain, memory, and wakefulness [3].
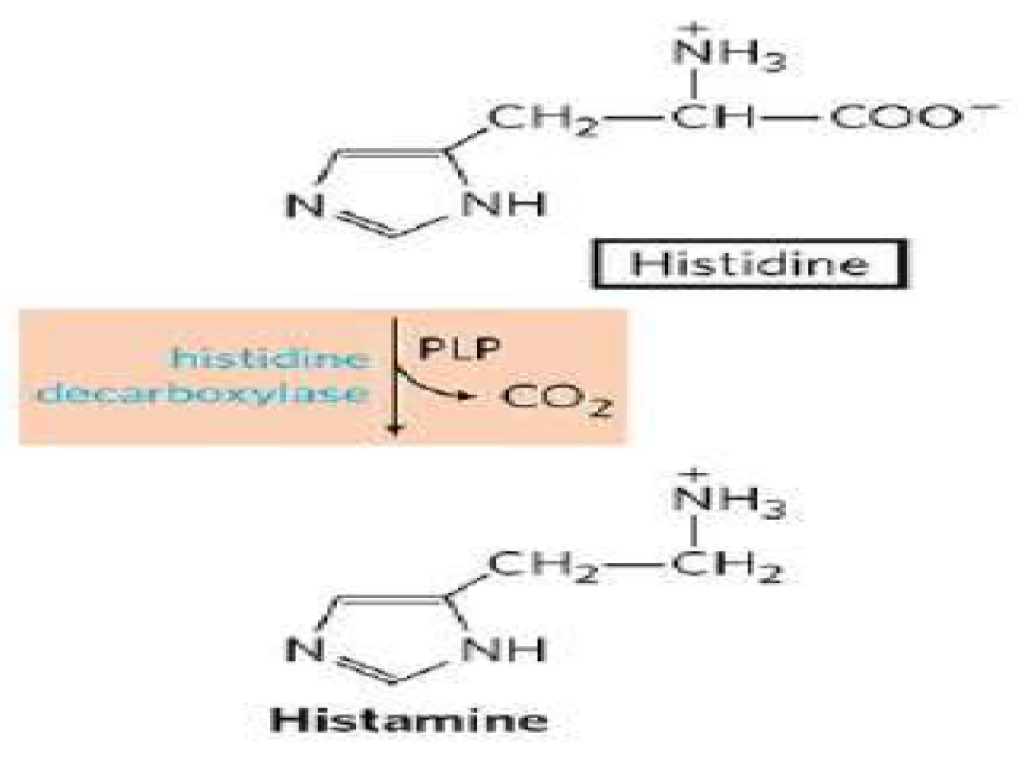
Figure 1: Synthesis of Histamine [4]
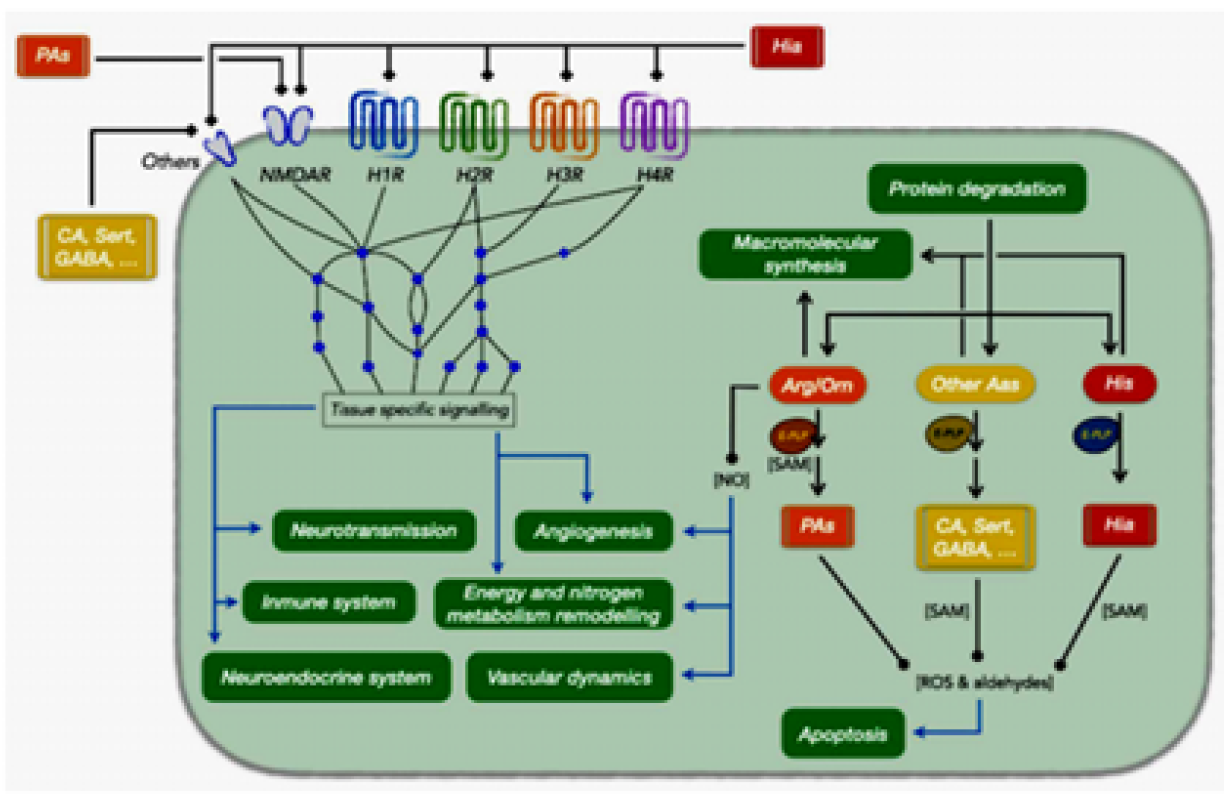
Figure 2: Relationships between Histamine and Other Amines with Different Physiological and Cellular Processes. The Processes are Depicted in Green Boxes. Abbreviations (by Alphabetical Order): Aas, Amino Acids; Arg/Orn, Arginine/Ornithine; CA, Catecholamines; E-PLP, Pyridoxal Phosphate Dependent Enzyme; GABA, Gamma Aminobutyric Acid; H1R, Histamine Receptor 1; H2R, Histamine Receptor 2; H3R, Histamine Receptor 3; H4R, Histamine Receptor 4; Hia, Histamine; His, Histidine; NMDAR, N-Methyl-D-Aspartic Acid Receptor; NO, Nitric Oxide; Pas, Polyamines; ROS, Reactive Oxygen Species; SAM, S-Adenosyl Methionine; Sert, Serotonin [1]
Chemical Structure, Synthesis and Metabolism of Histamine
Histamine is synthesized from the L‐histidine amino acid, by oxidative decarboxylation catalyzed by histidine decarboxylase enzyme (Figure. 1) which expressed in both the neuronal cell bodies and axon terminals, and the availability of L-histidine is the rate-limiting factor. Histamine is stored in vesicles in axon varicosities and cell bodies and by the vesicular monoamine transporter 2 and is released by exocytosis upon the arrival of action potentials. The release and synthesis of histamine are regulated by H3 receptors. In the brain, histamine is also produced by mast cells, which contribute modestly to the total amine levels in the adult brain [4]. Whether histamine is catabolized by diamine oxidase (DAO) or HNMT is supposed to depend on the localization of histamine. The DAO protein is stored in plasma membrane–associated vesicular structures in epithelial cells and is secreted into the circulation on stimulation. Therefore, it has been proposed that DAO may be responsible for scavenging extracellular histamine after mediator release. Conversely, HNMT, the second most important enzyme inactivating histamine, is a cytosolic protein, which can convert histamine only in the intracellular space of cells [4].
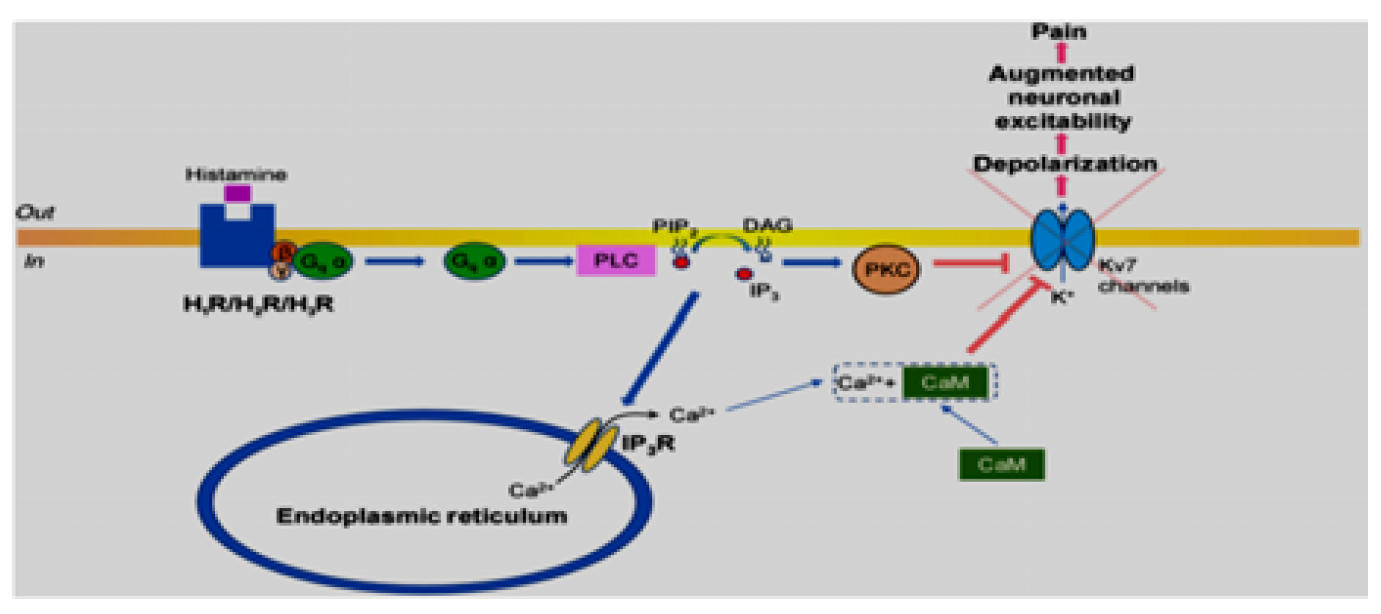
Figure 3: Diagram Illustrating Histamine Receptor Signaling-Gq pathway [3]
Histamine receptors
Histamine is a key player in allergic reactions, circadian cycle regulation, vascular permeability, gastric and neurological functions, immune cell differentiation, epithelium proliferation, neoplastic progression and angiogenesis. (Figure 1) [1].
The action of histamine is modulated by four types of G protein coupled receptors. These receptors are H1, H2, H3, and H4 which are different in their mode of signal transduction and pharmacological and physiological effects Figure 2.
H1 Receptor
H1 receptors are G protein coupled receptors causing excitation when combined to histamine ligand by activation of phospholipase C enzyme, which causes hydrolysis of phosphatidylinositol 4,5‐bisphosphate (PIP2) to produce diacyl glycerol (DAG) and inositol triphosphate (IP3). Diacyl glycerol activates protein kinase C and IP3 causes Ca2+ releasing from smooth endoplasmic reticulum and increase in cytoplasmic calcium ion concentration. This increased concentration of calcium ions activates phosphokinase C-‐dependent phosphorylation to form calcium-calmodulin complex that close voltage gated channel of potassium (Kv7) leading to depolarization and increased nociception [3] Figure 3.
H2 Receptor
H2 receptors in nervous system are postsynaptic, formation of ligation with histamine causes activation of adenylate cyclase enzyme causing splitting of ATP into cAMP by the action of Gs α subunit which energized by GPT. cAMP acts as a second messenger for activation of protein kinase A. Elevated cAMP concentration up‐regulates PKA, which as reported for hippocampal neurons, could activate ligand gated α‐AMPA receptors with resultant depolarization and increase of neuronal firing. Also, as it was reported for afterhyperpolarization in enteric neurons, PKA inhibits small conductance Ca2+‐activated potassium (KCa) channels with resultant augmented neuronal excitability [5] Figure 4.
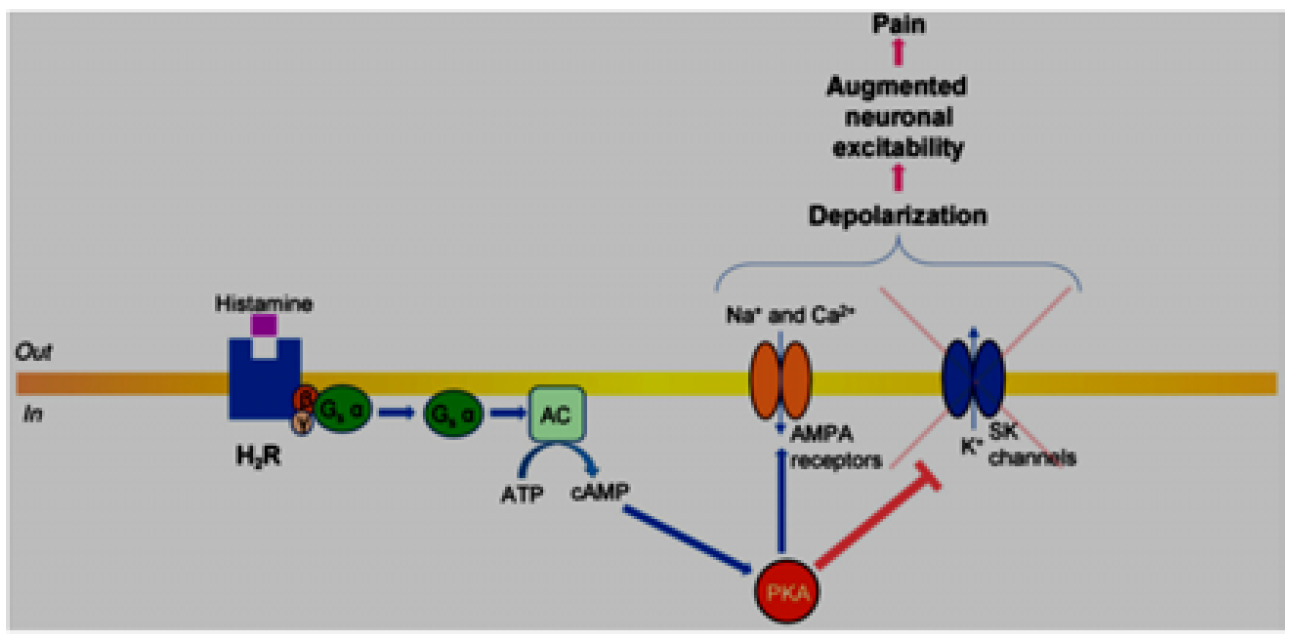
Figure 4: Diagram illustrating histamine receptor signalling—Gs pathway [3]
H3 Receptor
This type of histamine receptor is presynaptic receptor and coupled with Gi causing inhibition of adenylate cyclase by Gi α subunit and decreases cAMP production and protein kinase A activity. In sympathetic and sensory neurons, it was reported that H3 receptor activation stimulated dissociation of Gi β and γ subunits from Gi α subunit, which leads to closer voltage‐gated calcium ion channels and preventing Ca2+ entrance through N‐, P‐, and Q‐type Ca2+ channels, and stimulated G protein‐coupled inwardly rectifying potassium channels these mechanisms brings the membrane of presynaptic neurons to more negative and become hyperpolarized causing reduction of its excitability and relieving pain. Besides the Gi pathway, postsynaptic activation of H3 receptors was reported to stimulate phospholipase C enzyme in a subpopulation of striatal neurons, with subsequent activation of the IP3 pathway followed by increased intracellular concentrations of Ca2+. Thus, it is analogous to the mechanisms described for H1 /H2 receptors coupled to Gq proteins. Similarly, the H1 and H2 receptor Gq cascade PLC signalling pathways modulate neuronal excitability with resultant potential facilitation of pain sensitivity. Furthermore, H3 receptor activation was established to inhibit glycogen synthase kinase 3β (GSK3β) and MAPK/ERK pathways. These effects are translated via Gi β and γ subunits, which up‐regulate the PI3K pathway with subsequent production of phosphatidylinositol 3,4,5‐ triphosphate (PIP3) from PIP2, which results in the recruitment of PKB (Akt). PKB is initially activated by phosphoinositide‐dependent kinase 1 (PDK1) and mammalian target of rapamycin complex 2‐dependent phosphorylation. PKB, via phosphorylation, inactivates GSK3β, which decreases neuronal excitability, inhibits neuronal inflammation, and, subsequently, relieves pain. PKBdependent phosphorylation that activates the MAPK/ERK cascade was also reported to be a highly efficient neuroprotective mechanism for chronic inflammatory and neuropathic pain [3,6] Figure 5.
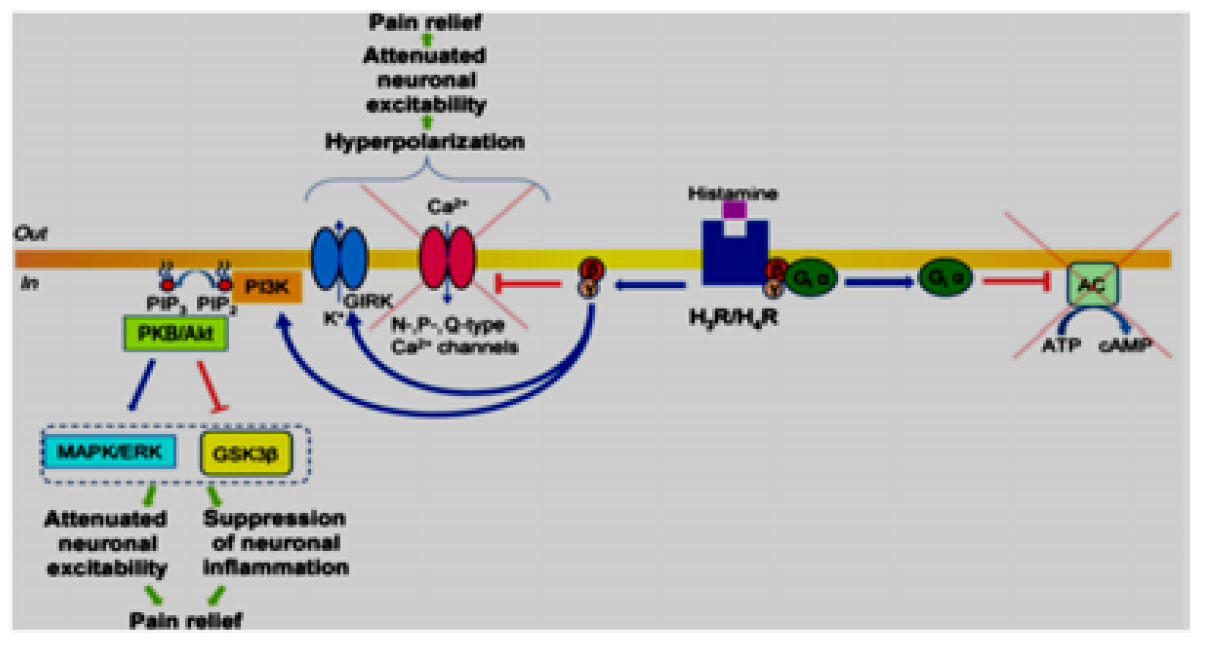
Figure 5: Diagram Illustrating Histamine Receptor Signalling-Gi Pathway [3]
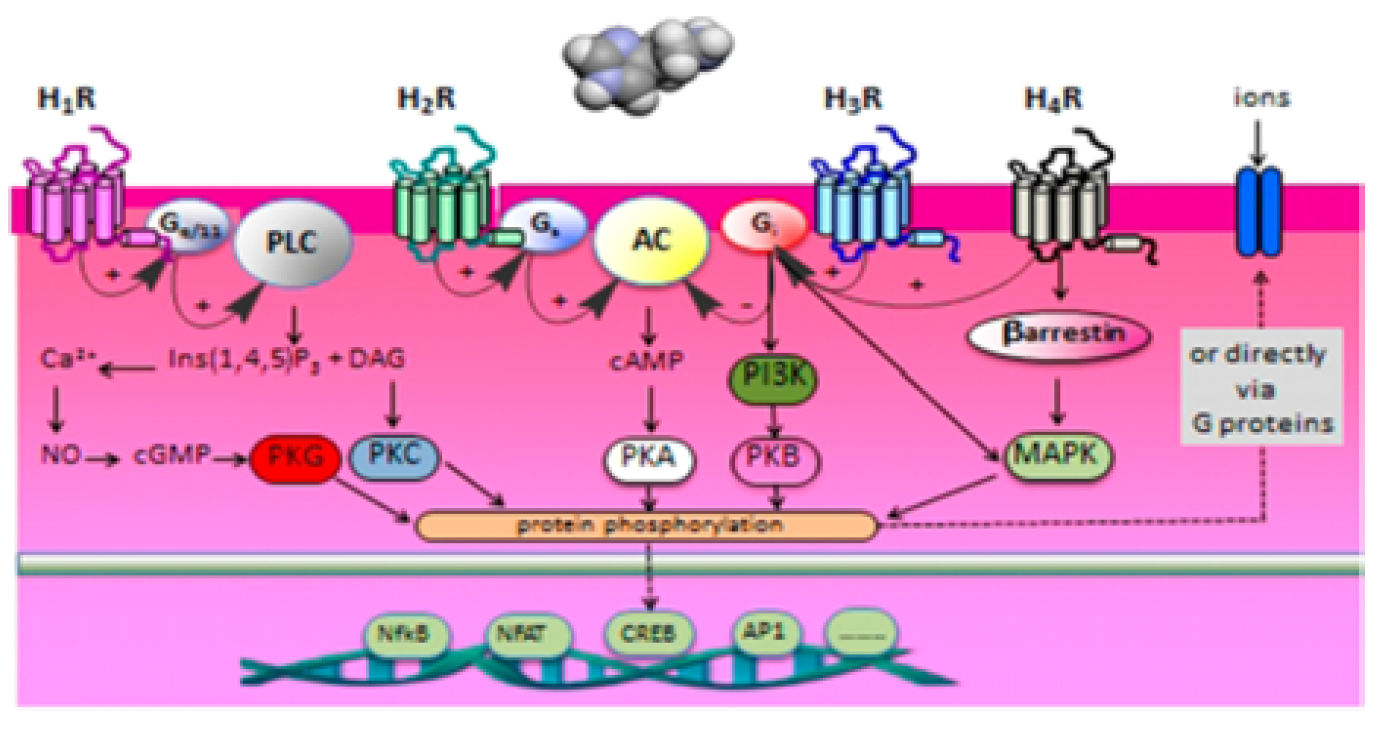
Figure 6: Schematic Overview of the Main Signal Transduction Routes of the four Different Histamine Receptors [2]
H4 Receptor
H4 receptors are known to be coupled to Gi proteins, and their downstream pathways are postulated to be similar to those described for H3 receptors. Compared to the other three types of histamine receptors, the H4 receptor is not expressed abundantly in the CNS and PNS. By quantitative single‐cell Ca2+ imaging, it was demonstrated that histamine induces a Ca2+ increase in a subset of sensory neurons (3–10%) via activation of the H1 and H4 receptors as well as inhibition of the H3 receptor. It is assumed that the decreased threshold in response to H3 receptor antagonism, which accounts for the analgesic effect of H3 receptor antagonists, activates H1 and H4 receptors on sensory neurons, which in turn results in the excitation of histamine‐sensitive afferents and, therefore, may result in a modulation of pain sensitivity [3] Figure 6.
Histamine as a Neurotransmitter
The histamine releasing and synthesizing neurons are found in tuberomamillary nucleus (TMN) in hypothalamus, these neurons send projections to entire CNS through ascending bundles to forebrain and descending one to spinal cord. The resting membrane potential for histaminergic neurons is 250 mv and spontaneously fire action potentials at 2.1 6 0.6 Hz and has circadian rhythmicity, which means that the higher activity noticed during Wakefulness periods. The histaminergic neurons activity and the behavioral state linked to it is influenced by several neuronal, humoral, and Paracrine, endocrine, and humoral signals, and the activity is regulated mainly by excitatory effect of glutamate releasing neurons coming from cerebral cortex and the hypothalamus and by inhibitory gama amino butyric acid releasing neurons from ventrolateral preoptic nucleus in hypothalamic. The synthesis of histamine L-histidine is catalyzed by HDC enzyme, which is expressed in both the neuronal cell bodies and axon terminals, and the availability of L-histidine is the rate-limiting factor. Histamine is stored in vesicles in axon varicosities and cell bodies and by the vesicular monoamine transporter 2 (VMAT-2) and is released by exocytosis upon the arrival of action potentials. The release and synthesis of histamine are regulated by H3 receptors. Most histaminergic fibers do not make typical synaptic contacts, and the amine is released from several points along the fibers, allowing its action on a large number of cells. In the brain, histamine is also produced by mast cells, which contribute modestly to the total amine levels in the adult brain [4].
Inactivation of histamine in the brain is due primarily to the action of histamine-N-methyltransferase producing telemethylhistamine that is transformed to telemethylimidazolacetic acid by monoamine oxidase B. Diamine oxidase is the main histamine-metabolizing enzyme in the peripheral tissues, but its activity in the brain is considerably lower under basal conditions. In contrast to most other aminergic neuronal cells, histaminergic neurons lack a specific reuptake transporter, although astrocytes take up histamine with low affinity (Km 0.56 mM and 4.0 mM) through the activity of the plasma membrane monoamine transporter and, to a lesser extent, the organic cation transporter 3 [7].
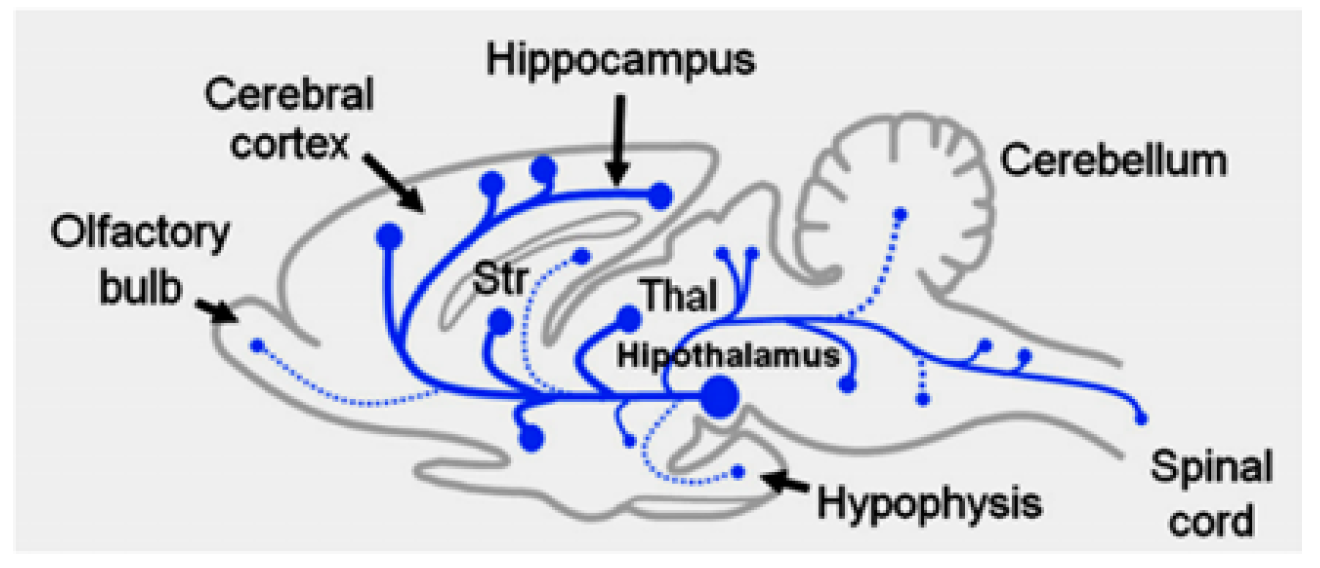
Figure 7: The Histaminergic System in the Rat Brain [7]
Action of Histamine in Cardiovascular System
Vasodilator effect of histamine on arterioles is well known, like the effect of bradykinin. It also increases the porosity of capillaries to leak plasma and plasma proteins into interstitial space. In inflammatory responses and allergic reactions, the vasodilator effect of histamine on arterioles and increase in capillary porosity leads to edema [8] Figure 8.
In spite of the low degree of sensation in cardiac muscle, in case of ischemia, the pain sensation is caused by tissue damage and release of histamine, bradykinin and other kinins, accumulation of acids and proteolytic enzymes and slow moving blood in coronary circulation which decrease washing of these substances from ischemic myocardium. These substances cause stimulation of nerve ending in myocardium, sending pain sensation signals to the central nervous system [8].
Release of toxic substance from ischemic tissue as histamine, tissue enzyme, and serotine is the main cause of circulatory shock. Anaphylactic shock mechanism caused by antigen-antibody reaction which occurs by entering antigen to circulation and causing basophils and mast cells in blood and tissue respectively to degranulate and release large quantity of histamine. Histamine effects are: increase in venous dilation and decrease of venous filling pressure and venous return, decrease of arterial pressure due to arteriolar dilation, rapid loss of plasma and plasma proteins to tissue due to increase of capillaries porosity. All of these mechanisms causes the anaphylactic shock [8].
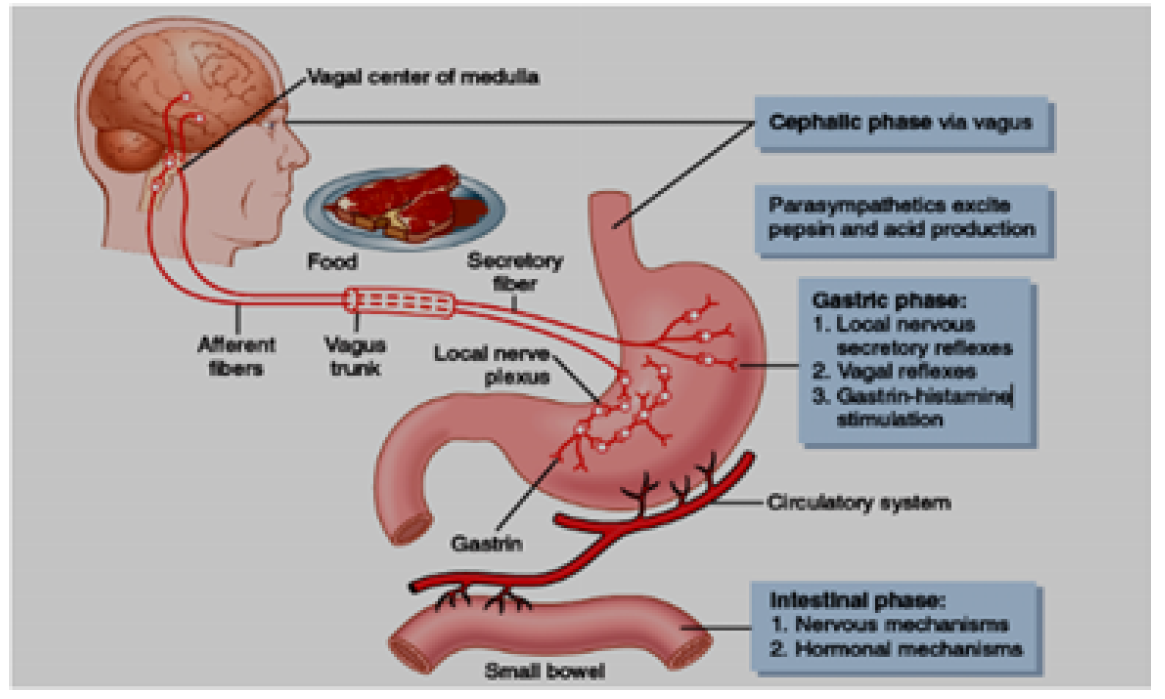
Figure 8: Phases of Gastric Secretion and their Regulation [8]
Action of Histamine on Gastric Secretions
Gastric secretion for hydrochloric acid, proteolytic enzyme pepsin and mucus by parietal cells, peptic cells, and mucus cells stimulated by the effects of acetylcholine, histamine and gastrin, but the main effects of histamine and gastrin on parietal cells which produce hydrochloric acid. The release of histamine from enterochromaffin-like cells (ECL cells) stimulated by gastrin which secreted by G cells in enteroendocrine gland action of presence of protein in food. The quick action of hydrochloric acid secretion is caused mainly by histamine. The ECL cells lie in the deep recesses of the oxyntic glands and therefore release histamine in direct contact with the parietal cells of the glands. The rate of formation and secretion of hydrochloric acid by the parietal cells is directly related to the amount of histamine secreted by the ECL cells. In turn, the ECL cells are stimulated to secrete histamine by the hormone gastrin, which is formed almost entirely in the antral portion of the stomach mucosa in response to proteins in the foods being digested. The ECL cells may also be stimulated by hormones secreted by the enteric nervous system of the stomach wall [8].
Effects of Histamine on Bronchia and Bronchioles
Local secretary factors may cause bronchiolar constriction. Histamine and slow reactive substance of anaphylaxis are substance formed in lung and cause bronchial constriction. Both of these substances are released in the lung tissues during allergic reactions by tissue mast cells by the action of allergic substances as pollen grains, irritant chemicals or vapors. Histamine and slow reactive substance causes contraction of bronchiolar smooth muscle and constriction of air way as occur in bronchial asthma. The same irritants that cause parasympathetic constrictor reflexes of the airways as, sulfur dioxide, smoke, dust. The acidic elements in smog acts directly on the lung tissues to initiate local, non-nervous reactions that cause constriction of small airways [8].
In persons with asthma, when asthmatic patient exposed to or inhale any of the substance which sensitive to it as pollen grains, dust, or smog, the antibodies on mast cells surface in lung tissue, reacts with and attaches to this irritants forming complex between antigen and IgE, this complex formation causes mast cells degranulation and release of histamine, slow-reacting substance of anaphylaxis (which is a mixture of leukotrienes), bradykinin and eosinophilic chemotactic factor. The action of these mediators causes contraction of bronchiolar smooth muscle and spasm, secretion of thick mucus and formation of edema in their walls leading to air way obstruction [8].
Action of Histamine in Immunity
The activities of histamine as one of the most important inflammatory mediators by its action on blood vessels as vasodilator its chemoattractant properties is well known. The binding of histamine to its receptors on eosinophils (H4) favoring eosinophils migration from blood to the infected tissue and of expression of macrophage-1 antigen and ICAM-1 adhesion molecules. The binding of histamine to H4 receptors on mast cell membrane, causes intracellular calcium release and recruitment of mast cells into tissues. The activation of mast cells in infected tissue amplifies the inflammatory response and may favor the establishment of a chronic inflammatory response [9].
The inflammatory mediation of histamine in air way occurs by its binding to lung macrophage by H1 receptors on their surface and production of IL-6 and β-glucuronidase, lysosomal release and damage of epithelium were associated with basement membrane rupture. The contribution of histamine to the induction of airway inflammation is also due to its effect on nonimmune cells. In nasal fibroblasts, there is a dose-dependent increase in IL-6 production in response to histamine stimulation. This inflammatory mediator increases expression of phosphorylated p38, pERK, and pJNK and induces NF-κB activation [9].
In the nervous system, microglial activation is regulated by histamine in a dose-dependent manner, which leads to the production of proinflammatory cytokines, such as IL-6 and TNF-α. This activation is mediated via H1R, and H4R and is dependent on MAPK and PI3K/AKT cascades. In addition to inducing iNOS expression and NO production, histamine promotes the loss of mitochondrial membrane potential and the production of ROS in microglia by binding to these same receptors. Overall, the accumulation of these cytokines and proinflammatory molecules can be deleterious, leading to nerve damage [10].
Histamine also modulates the response of DCs. Stimulation of immature DCs induces expression of CD86, CD80, and MHC class II, increasing the efficiency of T cell activation. In the presence of histamine, DCs exhibit a higher production of IL-6, IL-8, and CCL2 as well as induced expression of IL-1β, CCL5, and CCL4. However, the H2R pathway promotes IL-10 production and inhibits IL-12 synthesis by immature DCs, favouring the development of a Th2 response profile. This modulation of cytokine production, suggests that histamine indirectly alters the Th1/Th2 balance through the stimulation of DCs. Th1 cells display higher H1R expression, and their binding to histamine promotes activation of Th1 responses, potentiating IFN-γ production. In contrast, Th1 and Th2 responses are inhibited by histamine stimulation via H2R. Histamine also alters the response of other subpopulations of lymphocytes. Activation of CD8+ T cells via H4R induces secretion of IL-16, a chemoattractant molecule for CD4+ cells such as monocytes and DCs. In addition, histamine stimuli induces IL-17 production in human Th17 cells, suggesting the contribution of this inflammatory mediator to the activation of lymphocytes present in skin lesions in atopic dermatitis and psoriasis [9].
Other evidence, indicates that histamine plays an important role in inflammatory skin diseases. When stimulated via H4R, the NK cells present in skin lesions, increase expression of the chemokines CCL3 and CCL4, favouring cell recruitment to injured tissue . As H4R knockout mice display a lower influx of inflammatory cells and less cell proliferation at the lesion sites, H4R is associated with the inflammatory response in atopic dermatitis. H4R is also involved in NK cell recruitment and induction of CCL17 production by DCs at lesion sites in murine models of atopic dermatitis, contributing to increased local inflammation . Furthermore, higher production of IL-6 and adhesion molecules, such as ICAM-1 and VCAM-1, occurs in endothelial cells stimulated by histamine, thereby favouring progression of the disease. In an experimental model, the absence of H1R reduced the development of atherosclerosis, whereas the absence of H2R exerted the opposite effect [9] (Table 1).
Histamine Receptors and Pain
The stimulation of histamine receptors increases the pain sensation in nociceptive pathways, while inhibition or blocking of these receptors causes antinociceptive and neuroprotective.Many subtypes of histaminic receptors are found in postsynaptic or presynaptic neurons. Presynaptic neuron receptors function as heteroreceptors or autoreceptors or providing either negative feedback or positive feedback regulation for releasing of neurotransmitter into the synaptic cleft. The resultant inhibitory or excitatory effect of histamine receptors depends on the action of the neurotransmitter and the subsequent intracellular signaling. The presence of histamine receptors in postsynaptic or presynaptic neurons is specific for each type of receptor. H1 and H2 receptors are mainly present in postsynaptic neurons. H3 and H4 histaminic receptors have higher affinity to histamine than H1 and H2 and are predominantly expressed in neurons. Expression of H3 and H4 receptors on the opposite sides of the synaptic cleft may contribute to their effects in neuropathic pain. In addition to its localization on postsynaptic or presynaptic neurons, the physiological effect of histamine is also determined by the type of G-protein coupled receptor and the subsequent intracellular signaling. The difference in underlying signaling pathways may directly determine the effect on pain perception produced by selective ligands, even when they act at the same histamine receptor [3,11].
Table 1: Immunological Functions Mediated by Histamine Receptors Signaling [9]
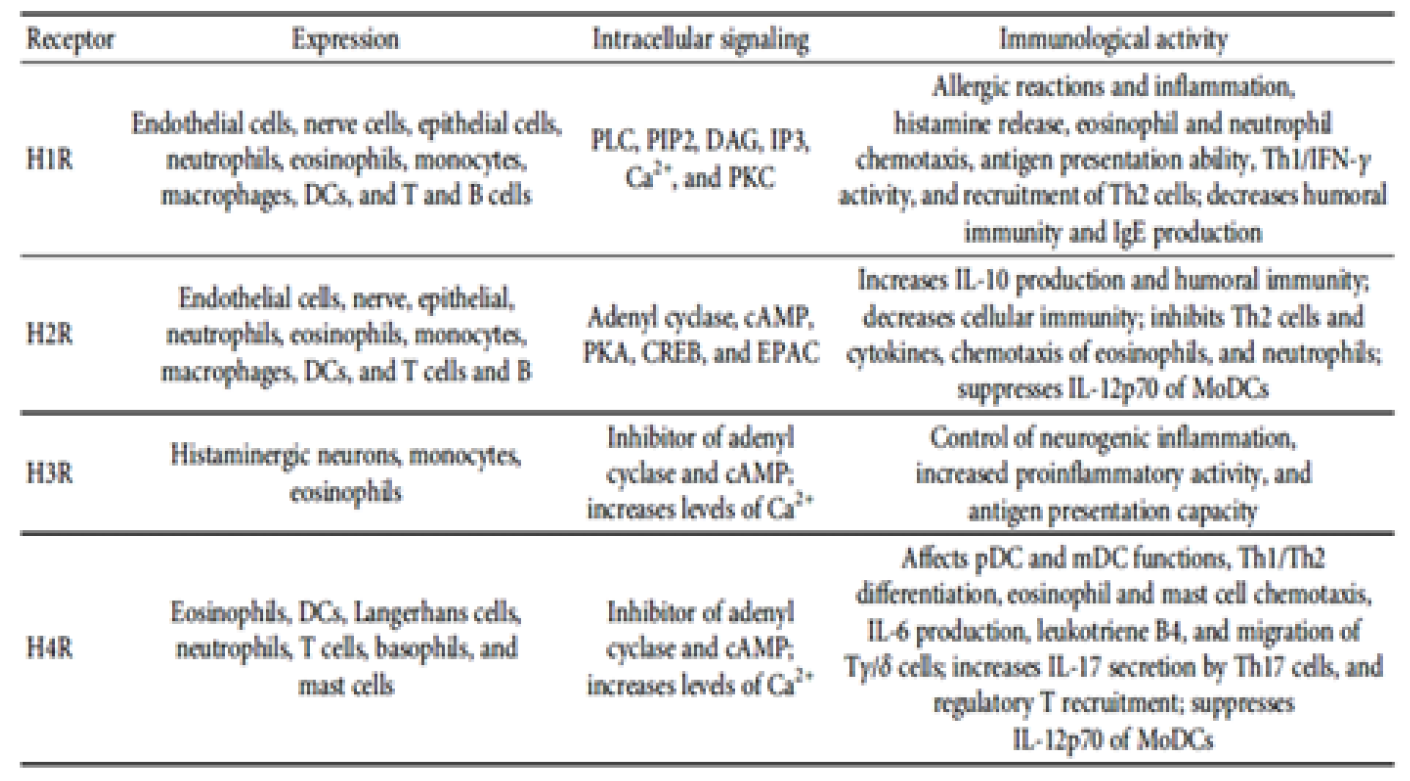
Histamine Intolerance
Histamine intolerance results from a disequilibrium of accumulated histamine and the capacity for histamine degradation. The main enzyme for metabolism of ingested histamine is diamine oxidase. An impaired histamine degradation based on a reduced DAO activity and the resulting excess of histamine may cause numerous symptoms mimicking an allergic reaction. Ingestion of histamine-rich food, alcohol, or drugs that release histamine or block DAO may provoke diarrhea, headache, congestion of the nose, asthmatoid wheezing, hypotension, arrhythmia, urticaria, pruritus, flushing, and other conditions in these patients. Approximately 1% of the population has histamine intolerance, and 80% of those patients are middle-aged. Because of the multifaceted symptoms, the existence of histamine intolerance is frequently underestimated, or its symptoms are misinterpreted [12].
It can be concluded that the action of histamine is modulated by H1, H2, H3, and H4 receptors. It is a key player in allergic reactions, circadian cycle regulation, vascular permeability, gastric and neurological functions, immune cell differentiation, epithelium proliferation, neoplastic progression and angiogenesis. The rate of formation and secretion of hydrochloric acid by the parietal cells is directly related to the amount of histamine secreted by the enterochromaffin-like cells. The release of histamine may be provoke diarrhea, congestion of the nose, headache, asthmatoid wheezing, hypotension, arrhythmia, urticaria, pruritus, and flushing.
Moya-García, A.A. et al. “Histamine, metabolic remodeling and angiogenesis: A systems level approach.” Biomolecules, vol. 11, 2021, article 415, pp. 1–16.
Panula, P. et al. “International Union of Basic and Clinical Pharmacology. XCVIII. Histamine receptors.” Pharmacological Reviews, vol. 67, 2015, pp. 601–655.
Obara, I. et al. “Histamine, histamine receptors, and neuropathic pain relief.” British Journal of Pharmacology, vol. 177, 2020, pp. 580–599.
Zimmermann, A.S. et al. “Systematic analysis of histamine and N-methylhistamine concentrations in organs from two common laboratory mouse strains: C57BL/6 and BALB/c.” Inflammation Research, vol. 60, 2011, pp. 1153–1159.
Park, P. et al. “Calcium-permeable AMPA receptors mediate the induction of the protein kinase A-dependent component of long-term potentiation in the hippocampus.” Journal of Neuroscience, vol. 36, 2016, pp. 622–631.
Schlicker, E. et al. “Role of the histamine H3 receptor in the central nervous system.” Histamine and Histamine Receptors in Health and Disease, vol. 241, 2016, pp. 277–299.
Nieto-Alamilla, G. et al. “The histamine H3 receptor: structure, pharmacology, and function.” Molecular Pharmacology, vol. 90, 2016, pp. 649–673.
Guyton, A.C. et al. Textbook of medical physiology. 13th ed., Saunders Elsevier, 2016.
Branco, A. et al. “Role of histamine in modulating the immune response and inflammation.” Mediators of Inflammation, 2018, article 9524075, pp. 1–10. https://doi.org/10.1155/2018/9524075
Rocha, S.M. et al. “Histamine: A new immunomodulatory player in the neuron–glia crosstalk.” Frontiers in Cellular Neuroscience, vol. 8, 2014, article 120.
Popiolek-Barczyk, K. et al. “Antinociceptive effects of novel histamine H3 and H4 receptor antagonists and their influence on morphine analgesia of neuropathic pain in the mouse.” British Journal of Pharmacology, vol. 175, 2018, pp. 2897–2910.
Maintz, L. et al. “Histamine and histamine intolerance.” American Journal of Clinical Nutrition, vol. 85, 2007, pp. 1185–1196.
