
+91 6002993949
submission@iarconsortium.org
Open Access
ISSN (Print) : 2788-9459
ISSN (Online) : 2788-9467
We reviewed the published information on the thyroid hormone system and the mathematical modelling associated with it. The important thyroid hormones, viz. T3 and T4, regulatory in various metabolic activities, are released as an outcome of complex interactions occurring in the hypothalamus, pituitary and thyroid gland via the synthesis and release of Thyrotropin-releasing hormone, Thyroid-Stimulating Hormone, and thyroid hormones, respectively. Various mathematical models had been proposed in the past to explain the function and regulation of these hormones. The models that interacted pituitary-thyroid systems provide a better understanding of the sensitivity of the pituitary to the feedback effect of thyroid hormones in the context of thyroid hormone excess and deficiency A computational thyroid model may capture the known aspects of thyroid physiology. These models also explain the mental disorder called periodic relapsing catatonia. Certain models used Langmuir adsorption isotherm to understand endocrinological homeostasis. Thus, the study of these models will help better understand the physiology and functions of the endocrine system and hormones, particularly, those related to thyroid glands.
Modelling of physiological systems has aroused considerable interest over the past several decades. The physiological systems, particularly the endocrine systems, can preserve their internal balance through the actions of feedback mechanisms that connect them to technological feedback control systems. These are widely studied in engineering fields, as their regulation abides essentially to the same principles. The formidable complexity and a large number of interactions inherent within and among endocrine systems introduce problems of quantification that well fit the tremendous abilities of computers, and that (verbal) language mostly fails to suffice, whereas modelling and simulation succeed for both simple and complex systems, as pointed out in pioneer work on the simulation of thyroid hormone dynamics [1].
Mathematical and computational modelling involve the translation of biological statements into equations or computational algorithms. Running these equations on a computer brings simulated or generated theoretical data that may be compared with observational data. This will help predict the outcome of biological interactions, particularly the human hormonal or endocrinological system. Hence, we reviewed the published literature on the endocrine system and the proposed mathematical models to predict the outcome of the functioning of this system and related endocrine glands.
Endocrinological System
Hypothalamus Sector: Hypothalamus, a key homeostasis regulator, is a small region of the brain located above the brain stem, which is the central element in the regulation of endocrine function due to its connections with the pituitary gland [2]. It synthesizes and secretes unique releasing and inhibitory hormones and factors, which coordinate the production and secretion of hormones from the anterior lobe of pituitary gland. It is about 4 grams in weight in adult humans [3]. It secretes certain hormones, which also include TRH or Thyrotropin-releasing hormone, which is synthesized and secreted by the parvicellular neurons of the paraventricular cells (PVNparv) and the periventricular nucleus (PeriVN). TRH, principally stimulates the synthesis and release of TSH (Thyroid-Stimulating Hormone). The connection between the hormones of the hypothalamus and the anterior pituitary is enabled via minute blood vessels called hypothalamic-hypophysial portal vessels. Through these portal vessels, TRH is transported to the anterior pituitary to trigger the secretion of TSH. The rate of TRH secretion is mainly determined by the level of free thyroid hormone levels in blood. Some portion of free T3 and T4 molecules impinges upon hypothalamic cells and couples with the receptors on these cells [3]. The amount of thyroid hormone-receptor complexes is the main determinant of the rate of TRH secretion. As aforementioned, the levels of thyroid hormones in blood negatively affect TRH secretion. So, as the amount of thyroid hormone-receptor complexes increase on the cells of the hypothalamus, the TRH output will decrease, and vice versa. In short, the rate of TRH secretion is inversely proportional to the number of thyroid hormones in the blood. Two factors affect the concentration of a hormone in blood; secretion of that hormone and rate of removal from blood. As most other hormones do, TRH is cleared from the body with a certain half-life, where half-life is the time it takes for half of the amount of a hormone to be cleared from the blood in our context. TRH has a half-life of 6.2 minutes.
In short term, changes in TRH secretion rate occur as the levels of thyroid hormones in blood dictate. However, there might be cases where the stimulation persists at far below or far above the baseline values. Relying upon the fact that a hormone, which provokes or inhibits the activity of a gland, can also affect its weight over the long term in certain cases, thyroid hormones can also influence the weight of the hypothalamus. There is no direct evidence that the weight of the TRH-secreting section of the hypothalamus can be altered according to the circulating thyroid hormone levels. However, there is evidence that the number of cells that secrete CRH, which is a hypothalamic hormone analogous to TRH in the regulation of hypothalamus-pituitary-adrenal (HPA) axis, substantially decline in subjects who externally receive the hormones that inhibit its secretion [4]. Extrapolating all this information to our case, the weight of the hypothalamus is taken as a variable quantity.
First, if the magnitude of stimuli from thyroid hormones is persistently far above the standard levels, it means that the secretory capacity of hypothalamus is consistently underutilized. In such cases, the specific portions of hypothalamus, which are in charge of the TRH secretion, would shrink not to retain the redundant capacity in vain. Second, if thyroid hormones constantly circulate at considerably below normal concentrations, i.e. if hypothalamus is persistently under-stimulated, then the hypothalamus would continually operate at above-normal levels, and thereby expand to adjust its capacity. So, it adjusts its capacity in the direction that the current needs of the body necessitate.
Pituitary Sector
The pituitary gland, or hypophysis, is a complex endocrine organ positioned in the sella turcica, a bony cavity at the base of the brain, and is linked to the hypothalamus by a stalk. The weight of this gland is approximately 600 milligrams in the adult humans. The pituitary secretes numerous hormones, which take part in various physiological processes by either acting directly on the target cells, or stimulating other endocrine glands to secrete hormones leading to alterations in body function. The human pituitary is comprised of two morphologically and functionally distinct glands that are connected to the hypothalamus, which are neurohypophysis and the adenohypophysis, also known as the anterior pituitary. The anterior lobe of pituitary comprises 75% of its gland, and its cells secrete six hormones. TSH is one of the hormones that the anterior pituitary is in charge of synthesizing and secreting, and the cells specialized for TSH are called thyrotrophs. The thyrotrophs, i.e. TSH-secreting cells, compose 5% of the anterior pituitary cells.
TSH is the principal regulator of thyroid hormone synthesis and secretion for it is the eventual messenger in the stimulation of the thyroid gland. As noted earlier, TRH plays a major role in the positive regulation of TSH secretion. Upon secretion, TRH reaches the anterior pituitary through the portal blood system, impinges upon the thyrotrophs, and binds to specific receptors on these cells. The binding of TRH with its receptors on thyrotrophs activates numerous intracellular mechanisms which ultimately lead to TSH release. However, it is not only TRH that influences the rate of TSH secretion, but also the levels of thyroid hormones in the blood. Thyroid hormones exert a suppressive, negative feedback effect on the thyrotrophs to prevent the oversecretion of TSH, as opposed to the augmenting effect of TRH. Some portion of circulating free thyroid hormones binds with the unique thyroid hormone receptors, TR’s, on the thyrotrophs, and exerts a suppressive effect on TSH release. This means that an increase in circulating thyroid hormone concentrations would lead to a reduction in the rate of TSH secretion; and a decrease would result in a rise in TSH secretion. Consequently, the magnitude of TSH secretion is induced by the opposing signals to the anterior pituitary, one by TRH and the other by the thyroid hormones [3]. The time it takes for the contrasting effects of TRH and thyroid hormones on TSH release to be revealed are different indeed. TRH elicits a prompt release of TSH within minutes (~15 minutes), while the inhibitory effect of thyroid hormones becomes evident after several hours [3]. Still, the time lag between the effect of TRH and thyroid hormones on TSH does not make much difference because the time unit of our model is one day and all these occur within a day anyway. Upon stimulation of the anterior pituitary by TRH, TSH is released into the circulation. TSH is typically measured in “microunits” (μU) or “milliunits” (mU). The normal range for TSH secretion rate is 40-150 μU/day, and for circulating TSH in plasma 0.3-4 μU/ml [5]. And, the half-life of TSH is about one hour.
It is stated that long-standing hypothyroidism may lead to pituitary enlargement, and increasing the negative feedback by any mechanism may result in atrophy of the thyrotrophs. In other words, prolonged overstimulation may lead to expansion in pituitary size and conversely sustained understimulation to shrinkage.
Thyroid Sector
Thyroid hormones are the primary regulators of metabolic functions in the body, and almost all the cells are regularly subjected to the actions of thyroid hormones. They are vital for the proper development and differentiation of the cells of the human body, and their deficiency results in serious or even life-threatening diseases. The human thyroid gland, located just below the larynx is the largest endocrine gland comprising of two lobes attached to either side of and anterior to the trachea. It normally weighs about 20 grams and is composed of spherical follicles filled with a gel-like substance called colloid, and surrounded by follicular cells (alternatively called thyrocytes). The primary ingredient of colloid is a large protein called thyroglobulin, which is the site where the thyroid hormones are formed and stored.
The synthesis and secretion of thyroid hormones initiate upon the stimulation of the thyroid by TSH. The binding of TSH with TSH receptors on follicular cells triggers a series of intracellular functions in these cells to synthesize and release thyroid hormones (Figure 1). For the synthesis of thyroid hormones, iodide is indispensable. So, the first step in the formation of these hormones is the uptake of available iodide into follicular cells. There, iodide is converted into an active form called iodine, which is the constituent part of thyroid hormones. Then, iodine attaches to tyrosine residues within thyroglobulin molecules. This process, i.e. binding of iodine to tyrosine residues within thyroglobulin, is called organification. The coupling of one iodine atom with a tyrosine molecule creates monoiodotyrosine (MIT). When MIT is iodized once more, a diiodotyrosine (DIT) molecule is formed. So, tyrosine is first iodized to monoiodotyrosine and then to diiodotyrosine. And, thyroid hormones are formed from these two kinds of molecules. When one MIT and one DIT molecule couples, one molecule of triiodothyronine, T3, is formed; and when two DIT’s come together, one molecule of thyroxine, T4, is created. So, the formation of one unit of T4 necessitates double the amount of iodine used for the production of one unit of T3. The molecular weights of T3 and T4 are 651 and 777 Da, respectively [3], where Da (Dalton) is a unit commonly used to measure mass on atomic or molecular scale and has a value of about 1.66054 × 10-24 g. After being synthesized, thyroid hormones reside in thyroglobulins, unless cleaved from them on demand. Stimulation of thyroid glandular cells by TSH not only promotes the synthesis of thyroid hormones, but also the secretion. The most rapid impact of TSH stimulation on the thyroid is the initiation of the breakdown of thyroglobulin, which results in the secretion of T3 and T4 within half an hour. For this to happen, follicular cells engulf bits of the colloid, internalizes the colloid droplet into the cells, disintegrates it with the help of enzymes, allow the hormones to discharge from the storage element thyroglobulin, and release them into the blood circulation. So, TSH stimulation simultaneously activates all the secretory mechanisms of thyroid glandular cells, but the fastest among them is the secretion process. Though synthesis requires a longer time compared to the secretion process, it still happens within one day under normal conditions. One unique characteristic of the thyroid gland, in contrast to most endocrine glands, is that it has a considerably large capacity to store thyroid hormones in itself. Thyroid gland can store approximately two months’ supply of thyroid hormones. Thus, if the synthesis of thyroid hormone ceases, the physiologic effects may not be recognized for about two months. In a sense, these stores serve as buffers to guard against sudden and transient dysfunctions in the thyroid system and thus helps preserve the healthy state of the body.
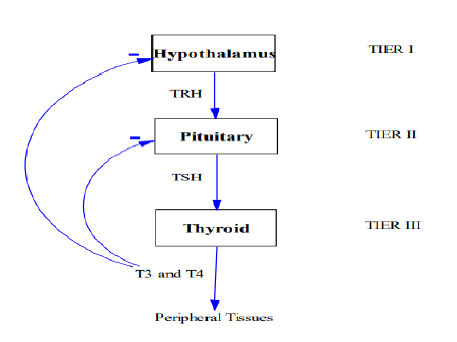
Figure 1: Diagrammatic Representation of Hormonal Flow Though Different Endocrine Glands
Normally, T4 constitutes 93% of the thyroid hormones released daily from the thyroid gland, and only 7% is T3. The functions of these two hormones are qualitatively the same; however, they differ from each other in terms of rapidity and intensity of action. T3 is said to be the physiologically active form of the thyroid hormones. T4 can only be synthesized in the thyroid gland and is the major secretory product of the thyroid gland. The normal production and secretion rate of T4 from the thyroid is approximately 90 μg/day. On the other hand, only 20% of T3 is produced directly by the thyroid; the rest is generated by enzymatic removal of one iodine atom (deiodination) of T4 molecules in peripheral tissues. The production rate of T3, including the peripheral conversions, is 35 μg/day [3]. Upon stimulation by TSH, T3 and T4 are released into the bloodstream. Most of the T3 and T4 molecules become bound to plasma proteins, only less than 1% of them circulate in free form. Normally, the range for total plasma concentration is 4-11 μg/dl for T4, and 75-220 ng/dl for T3. Specifically, 0.02% of T4 and 0.3% of T3 circulates in an unbound form. Merely the free portion of the hormones is biologically active and able to interact with target cells. And so, it is the circulating free hormones that feedback to the hypothalamus and pituitary to reduce the secretion of TRH and TSH [3].
In general, hormones are “cleared” from the plasma by various means involving metabolic destruction by the tissues, binding with the tissues, excretion by the liver into the bile, and excretion by the kidneys into the urine. For our case, it was recognized early that T4 and T3 were dispersed widely into tissues in addition to the blood. It is stated that about 40% of plasma T4 is converted to T3 and about 40% to reverse triiodothyronine (rT3), which is a metabolically inactive form. The half-life of T4 in the bloodstream is approximately 7 days, whereas that of circulating T3 is about 1 day. Like the hypothalamus and pituitary, the weight of the thyroid gland may also be altered under certain circumstances. The work of Vassart and Dumont [6] asserts that hypophysectomy, hypopituitarism, or an isolated TSH deficiency, i.e. the conditions that abate the thyroid function, leads to thyroid atrophy. Conversely, chronic stimulation of the thyroid for some reason is stated to enhance thyroid growth.
Modelling Endocrine System
Certain studies focused on the engineering principles that are applied to model the thyroid hormone system at various levels of complexities. One prominent name, in these studies, is Joseph J. DiStefano III. Numerous pioneering researches conducted by DiStefano and colleagues to model the thyroid system with a systems-theoretical approach and integrate it with experimental data. Some of these studies deal with hypotheses about the underlying feedback structure [7], some with parameter estimation for thyroid hormone secretion, distribution, binding, conversion and metabolism [1], some with the mathematical models for secretory output of thyroid hormones in response to TSH input [7], while a few with the prescription of thyroid hormones in hypothyroidism and after thyroidectomy [8].
Other thyroid-related quantitative modelling studies also adopt a systemic perspective. A mathematical model that interacted pituitary-thyroid provides a better understanding of the sensitivity of the pituitary to the feedback effect of thyroid hormones in the context of thyroid hormone excess and deficiency [9]. Liu et al. [10] proposed a model for the secretory system of the hypothalamo-pituitary thyroid axis by revising and improving the two works of Liu and Peng [11] and Liu and Liu [12] which accounted for the interactions of the hormones in the axis and the binding characteristics of hormones to proteins in plasma and tissues. Lastly, a computational thyroid model captures the known aspects of thyroid physiology and uses it to evaluate the competing hypotheses related to the Wolff-Chaikoff escape [13].
Danziger and Elmergreen [14] developed a model based on thyroid and pituitary hormones to explain the mental disorder called periodic relapsing catatonia. They proposed a set of below-mentioned nonlinear differential equations using Langmuir adsorption isotherm.
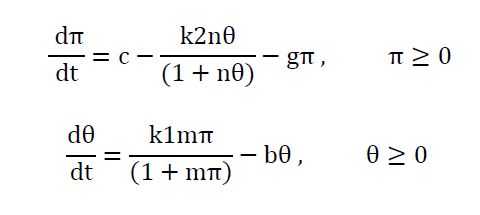
Where π and θ are concentrations of TSH and thyroid hormone respectively at time t and c, b, k1, k2, m and n are positive real constants. This describes most of the normal and malfunctions of the thyroid-pituitary system but the system fails to produce the sustained oscillations of the hormone levels, which was believed to be the reason for periodic relapsing catatonia. Danziger and Elmergreen [15] proposed a model that accounted for the sustained oscillations of the thyroid hormone levels, in addition, to explain the normal and abnormal operations of thyroid-pituitary system. They assumed that the pituitary gland secretes TSH, which activates an enzyme in the thyroid gland. The rate of production of thyroid hormone is considered to be proportional to the concentration of that enzyme. This mathematical model is as follows:
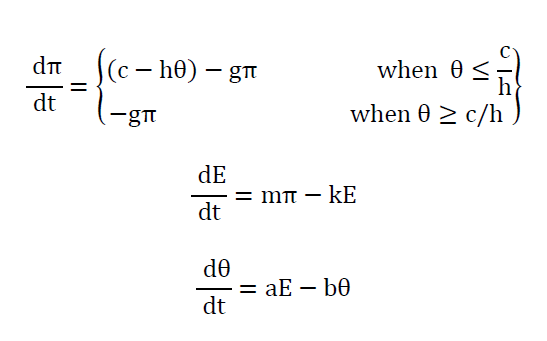
Where π, E and θ are concentrations of TSH, an enzyme and thyroid hormone at time t. The system is simply referred to as thyroid-pituitary regulator. There are two notable observations from this model apart from producing sustained oscillations, firstly they linearized the nonlinear terms and added a third differential equation and secondly, by varying specific model parameters, they explained the clinical conditions such as hyper - and hypothyroidism. Roston [16] presented a model of endocrinological homeostasis. The model had no enzymatic reaction terms and periodic solutions but had the autonomous secretion term for both TSH and thyroid hormones (T3 and T4) from the pituitary and the thyroid. In addition, he assumed i) thyroid hormones bound to serum proteins such as (thyroid-binding globulin (TBG) and albumin), ii) the physiological volumes Vp ,Vx in which TSH and thyroid hormones are dissolved are constant over a short period, and iii) the rate of secretion of thyroid hormones is proportional to the rate at which TSH passes through the thyroid gland. The model of (4) is as follows:
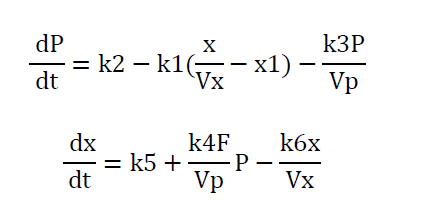
The notable features of this model are all the equations are stated in terms of amounts of TSH and thyroid hormones, which are distributed homogeneously and instantaneously throughout the physiological volumes Vp, Vx and the parameter k1 represents the sensitivity of the pituitary gland to thyroid hormones inhibition. Rashevsky [17] published a heterogeneous model of thyroid hormone regulator in a discussion of a mathematical theory of the effects of cell structure and diffusion processes on the homeostasis and kinetics of the endocrine system. The author kept all the basic assumptions of (3) and added the effects of the highly heterogeneous assumption of thyroid hormone regulator system. Norwich and Reiter [18] published a homogenous model of thyroid hormone regulation involving a set of linear differential equations expressing the relationship between the rates of secretion of thyroxine and of TSH. Using their model, they were able to replicate the known behavior of thyroxine and TSH and made certain predictions which are amenable to experimental verification or disproval by existing techniques. DiStefano and Stear [19] published a model of thyroid hormone regulation including the hypothalamus. Saratchandran et al. [20] published an improved mathematical model of thyroid hormone regulation by the anterior pituitary gland which accounted for experimental data of TSH and thyroid hormones. Degon et al. [12] published a computational model of the human thyroid gland based on the clinical observation of changes in the dietary iodine and the molecular pathways in the thyroid gland. This model captures the known aspects of thyroid physiology.
Pandiyan [21] presented a 4d model which was primarily aimed at the middle-age women or adult patient groups, since most of these patients regulate free T4 successfully for several years despite an increase in TSH and the presence of anti-thyroid antibodies (TPOAb and TGAb) in their blood serum
Their model was-
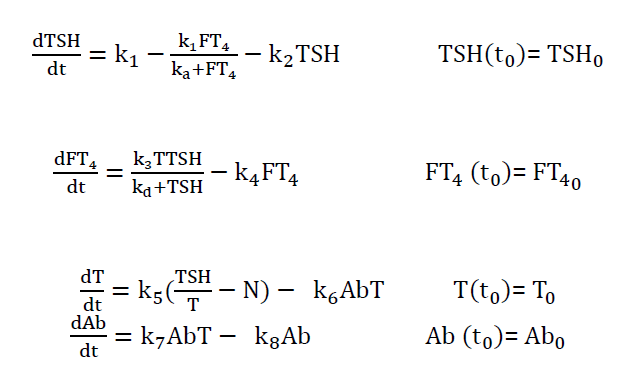
Hence, the above review supports the idea that the hormonal system can be simulated by appropriate mathematical models to better understand the functions related to endocrine glands.
DiStefano, J.J.3rd and R.F. Chang. “Computer simulation of thyroid hormone binding, distribution, and disposal dynamics in man.” American Journal of Physiology, vol. 221, no. 5, 1971, pp. 1529–1544. https://doi.org/10.1152/ ajplegacy.1971.221.5.1529.
Rhoades, R.A. and D.R. Bell. Medical physiology: Principles for clinical medicine. 3rd ed., Lippincott Williams & Wilkins, 2009.
Bhagavan, N.V. Medical biochemistry. 4th ed., Academic Press, 2002.
Erkut, Z.A. et al. “Glucocorticoids suppress corticotropin-releasing hormone and vasopressin expression in human hypothalamic neurons.” The Journal of Clinical Endocrinology & Metabolism, vol. 83, no. 6, 1998, pp. 2066–2073. https://doi.org/10.1210/jcem.83.6.4881.
Oertli, D. and R. Udelsman. Surgery of the thyroid and parathyroid glands. Springer-Verlag, 2007.
Vassart, G. and J.E. Dumont. “The thyrotropin receptor and the regulation of thyrocyte function and growth.” Endocrine Reviews, vol. 13, no. 3, 1992, pp. 596–611. https://doi.org/10.1210/edrv-13-3-596.
DiStefano, J.J.3rd. “A model of the regulation of circulating thyroxin unbound and bound to plasma proteins and its response to pregnancy, drugs, long-acting thyroid stimulator, and temperature stress.” Mathematical Biosciences, vol. 4, nos. 1–2, 1969, pp. 137–152.
Ben-Shachar, R. et al. “The biochemistry of acetaminophen hepatotoxicity and rescue: A mathematical model.” Theoretical Biology and Medical Modelling, vol. 9, 2012, p. 55. https://doi.org/10.1186/1742-4682-9-55.
Khee, M. and S. Leow. “A mathematical model of pituitary–thyroid interaction to provide an insight into the nature of the thyrotropin–thyroid hormone relationship.” Journal of Theoretical Biology, vol. 248, no. 2, 2007, pp. 275–287.
Liu, Y. et al. “A new mathematical model of hypothalamo-pituitary-thyroid axis.” Mathematical and Computer Modelling, vol. 19, no. 9, 1994, pp. 81–90.
Liu, B.Z. and J.H. Peng. “A mathematical model of hypothalamo-pituitary-thyroid axis.” Acta Biophysica Sinica, vol. 8, 1990, pp. 431–437.
Liu, Y.W. and B.Z. Liu. “An improved mathematical model of hypothalamo-pituitary-thyroid axis.” Progress in Biochemistry and Biophysics, vol. 6, 1992, pp. 439–443.
Degon, M. et al. “A computational model of the human thyroid.” Mathematical Biosciences, vol. 212, no. 1, 2008, pp. 22–53.
Danziger, L. and G. Elmergreen. “Mathematical theory of periodic relapsing catatonia.” Bulletin of Mathematical Biology, vol. 16, no. 1, 1954, pp. 15–21.
Danziger, L. and G. Elmergreen. “The thyroid-pituitary homeostatic mechanism.” Bulletin of Mathematical Biology, vol. 18, no. 1, 1956, pp. 1–13.
Roston, S. “Mathematical representation of some endocrinological systems.” Bulletin of Mathematical Biology, vol. 21, no. 3, 1959, pp. 271–282.
Rashevsky, N. “Mathematical theory of biological periodicities: Formulation of the n-body case.” Bulletin of Mathematical Biology, vol. 30, no. 4, 1968, pp. 735–749.
Norwich, K.H. and R. Reiter. “Homeostatic control of thyroxin concentration expressed by a set of linear differential equations.” Bulletin of Mathematical Biophysics, vol. 27, no. 2, 1965, pp. 133–144.
DiStefano, J.J.3rd and E.B. Stear. “On identification of hypothalamo-hypophysial control and feedback relationships with the thyroid gland.” Journal of Theoretical Biology, vol. 19, no. 1, 1968, pp. 29–50.
Saratchandran, P. et al. “An improved mathematical model of human thyroid hormone regulation.” Clinical Endocrinology, vol. 5, no. 5, 1976, pp. 473–483.
Pandiyan, B. Mathematical modeling and dynamical analysis of the operation of the hypothalamus-pituitary-thyroid (HPT) axis in autoimmune (Hashimoto’s) thyroiditis. 2011, Marquette University, Dissertation. http://epublications.marquette.edu/dissertations_mu/139.
