
+91 6002993949
submission@iarconsortium.org
Open Access
ISSN (Print) : 2708-5090
ISSN (Online) : 2708-5104
In the present study, detailed biology of Cavariella aegopodii has been studied during spring (March-April), summer (May-June), autumn (September-October) and winter (January-February) seasons under laboratory conditions to ascertain the pre-reproductive, reproductive and post-reproductive periods. Studies on the biology of Cavariell aaegopodii revealed that morphs of this species occurring on willow plants are parthenogenetic wingless female, parthenogenetic winged female, winged female sexuparae, oviparae and males. All the parthenogenetic wingless females undergo four regular moults resulting in five instar stages. In the winter season, the average nymphal period as well as pre-reproductive and reproductive periods is longer. The winged female sexuparae start appearing in apterous parthenogenetc colonies at the end of July month and their appearance continues upto November. Ovipara are apterous and males are winged.The present investigations were carried out in Shimla, Mashobra, Shoghi and Solan localities and as well as on plants maintained under laboratory conditions.
Aphids belong to insect superfamily Aphidoidea which evolved 280 million years ago. These tiny insect pests cause considerable damage to horticultural and agricultural crops by sucking the plant sap as well as by spreading a number of virus diseases (Kennedy et al., 1962). Many aphid species exhibit a life cycle in which parthenogenetic viviparous generations are followed by a single sexual generation and such species are called holocyclic. However, many aphid species have lost the sexual phase of their life cycle and are termed as anholocyclic. These tiny insect pests multiply rapidly through a series of parthenogenetic generations and in the sexual phase of their life cycle, genetic recombinations occur. The occurrence of polymorphism, viviparity and telescoping of generations in aphids make biological investigations in these insect pests very difficult but interesting (Blackman, 1980).The genus Salix, comprising of willow plants, is of great economic importance because of wide use of its species in a number of sectors (Kuzovkina and Quigley, 2005). Due to their pioneering characteristics and regenerative ability, many Salix species are of special importance for nature conservation and biological engineering (Holdenrieder et al., 1999).
Cavariellaaegopodii Scopoli have been reported as major pests of Salix spp. Cavariellaaegopodii, also known as carrot-willow aphid is wide spread throughout the temperate and warm temperate regions. This species is generally heteroecious and holocyclic in most parts of the world, however, in warmer climates, it is reported to be anholocyclic (Muller and Scholl, 1958; Dunn, 1965; Dunn and Kirkley, 1966). The primary host of Cavariellaaegopodii is Salix spp. and it migrates to secondary host such as carrots, celery and many other wild and cultivated species of Umbelliferae (Blackman and Eastop, 1994). Dunn (1965) reported that in Britain Cavariellaaegopodii Scopoli, goes through three parthenogenetic generations on the primary host (willow) in spring when the willow plants are actively growing and then migrates to secondary host (Umbelliferae plants) in summer, where they pass through several wingless and winged parthenogenetic generations. At the onset of autumn, these aphids produce first autumnal migrants and then males. The autumnal migrants return to the primary host and produce sexual females, which mature at the same time as the males arrive from the secondary host. The sexes mate and the overwintering eggs are laid on willow plants (Dunn, 1965).
In India, Cavariellaaegopodii has been reported from Northwestern and Northeastern Himalayas (Raychaudhuri, 1980). However, its detailed biology and cytogenetics have not been reported so far from India. Since the behavior and biology of this insect pest differ considerably in different regions due to different environmental conditions, it is desirable to study its biology and cytogenetics in Himachal Pradesh. Studies on the biology of Cavariellaaegopodii would be useful in ascertaining the occurrence of different morphs and understanding complex life cycles of these aphids in this region.
Such studies may prove useful in developing the effective strategies for the management of willow aphids.
Such studies may prove useful in developing the effective strategies for the management of willow aphids.
In the present investigations, regular surveys of different localities i.e. Shimla (31°1´ N Latitude, 77° 1´E Longitude), Mashobra (31°1´ N Latitude, 77° 2´E Longitude), Shogi (31°1´ N Latitude, 77° 5´ E Longitude) and Solan (30° 9´ N Latitude, 77° 15´E Longitude) were made every month during the years 2002 to 2007 in order to study the occurrence of different morphs of Cavariellaaegopodiiin natural population and observations were made on their biological characteristics. These aphid species were found infesting Salix babylonicatrees. Cavariellaaegopodii aphids were found in groups on leaves and stem.
Stem cuttings from older trees of Salix babylonica were taken and grown in pots (with a hole at the bottom) filled with loam soil. Fresh leaves appeared within 1 to 2 months time. These pots were placed in well protected chambers at room temperature. The adult apterous parthenogenetic viviparous females were taken from the mature infested Salix babylonica trees and were established on uninfestedSalix nursery plants grown in pots to obtain young nymphs. Once the nymph was laid by the adult female, the latter was removed from the plant. The nymphs thus obtained were kept under continuous observations till they got established on young nursery plants. Nymphs were allowed to grow, mature and produce the progeny on Salix babylonica plants.
Life history studies were made in the laboratory research chambers at room temperature. The detailed biology of Cavariellaaegopodiiwas studied during the whole year. The months of the year were grouped into seasons such as spring, summer, autumn and winter. Newly laid nymphs were transferred to new uninfested plants. A magnifying lens was used to follow the development and duration of different developmental stages. The total number of instars and duration of each instar stage were recorded. Data on the pre-reproductive, reproductive and post-reproductive periods of all the morphs of both the species occurring in this region were recorded. The temperature records of laboratory chambers from 2016 to 2019 where life history studies were conducted are given in the Table 1. The photographs of different morphs and their stages were taken with the help of digital still camera SONY cyber-shot DSC-W35 7.2 mega pixels.
The Cavariellaaegopodii aphids are generally found clustered together on leaves and stems of willow plants. Most of the aphids preferred to feed on young succulent growing parts. The aphid colonies produce large amount of honey dew. Studies on the biology of Cavariellaaegopodii revealed that morphs of this species occurring on willow plants are parthenogenetic wingless female, parthenogenetic winged female, winged female sexuparae, oviparae and males (Fig. 1).
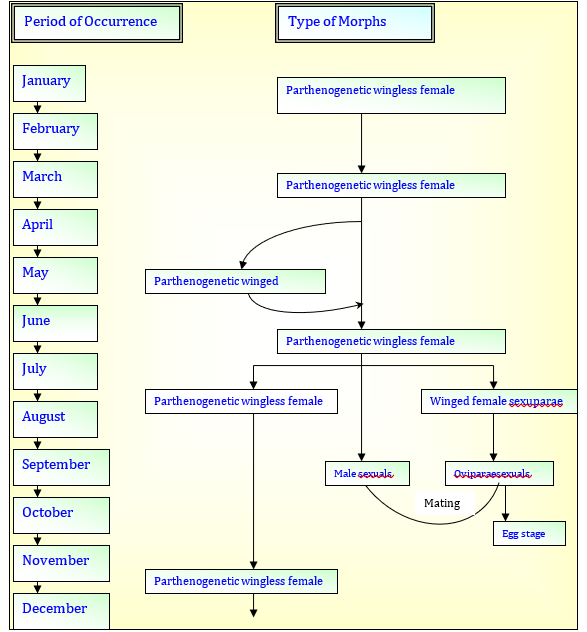
Fig. 1 Seasonal occurrence of different morphs of Cavariellaaegopodii on Salixbabylonica plants.
The biological characteristics of these morphs and duration of different stages are as follows
MORPHS OF CAVARIELLA AEGOPODII
Parthenogenetic wingless female
Parthenogenetic wingless females occur throughout the year. The newly laid nymph (Fig. 2) is pale yellowish green. The first instar nymph is not active after birth but later this moves to establish on softer part of the plant. Newly laid nymphs, prefer to stay along mid rib of leaves while those on stems generally face in downward direction. The second instar nymph (Fig. 3) is pale yellowish green in colour. Second instar nymphs are more active in comparison to first instar nymphs. The third instar (Fig. 4) is also yellowish-green in colour. Eyes are quite prominent and black in colour in this instar. Body is elongated with swollen abdomen, which has developing embryos. The fourth instar (Fig. 5) nymph is yellowish green in colour. It has elongatesiphunculi. The adult aphid (Fig. 6) is yellowish green with siphunculi usually tubular stout, slightly to moderately swollen distally. Legs and siphunculi are mainly pale. Backwardly directed supracaudal process bears a pair of hairs near apex.

Parthenogenetic winged female
Parthenogenetic winged females occur during summer season (May-June) only. Large colonies of aphids results in production of parthenogenetic winged females amongst the parthenogenetic wingless colonies. The newly laid nymph (Fig. 7) is pale yellowish green. The first instar nymphs move to establish on softer part of the plant. The second instar nymph (Fig. 8) is pale yellowish green in colour and is more active in comparison to first instar nymph. The third (Fig. 9) and fourth instar (Fig. 10) nymphs are yellowish green in colour and have elongated siphunculi. The adult aphid (Fig. 11) is yellowish green with siphunculi usually tubular stout, slightly to moderately swollen distally. These winged females were found to disperse to new plants to cause new infestation. Body of this morph is elongated and dull yellowish green in colour. Siphunculi are long, tubular and swollen at distal end.

Winged female sexupara
At the end of July month, the alate female sexuparae start appearing in apterous parthenogenetc colonies and their appearance continues upto November. The winged female sexuparae give birth to ovipara sexuals only. Female sexuparae also undergo four moults resulting in five instar stages. The first instar nymphs (Fig. 12) of winged female sexuparae are yellowish green in colour. Body is slightly elongated and wing pads are not visible in first instar stage. The colour of second instar (Fig. 13) is yellowish brown in colour. The third instar (Fig. 14) is dark yellowish brown in colour. Eyes are large and black in colour. The wing pads are quite prominent. The fourth instar (Fig. 15) is yellowish brown in colour and has elongated body. Dorsum of abdomen is pigmented with a median brownish patch on segments 2 to 6, which may sometimes be broken and segments seven and eight have distinct transverse bands. Siphunculi are longer and reach upto cauda which is conical. Subanal plate is protruded having semioval posterior margin. Subgenital plate has 8 to 10 hairs. The head and thorax of adult alate female sexuparae (Fig. 16) are dark black in colour and abdomen is yellowish brown in colour. Rostrum of adult extends upto middle coxae. The antennae are six segmented. Siphunculi of this morph are also long, with slightly swollen tips.

Ovipara sexual
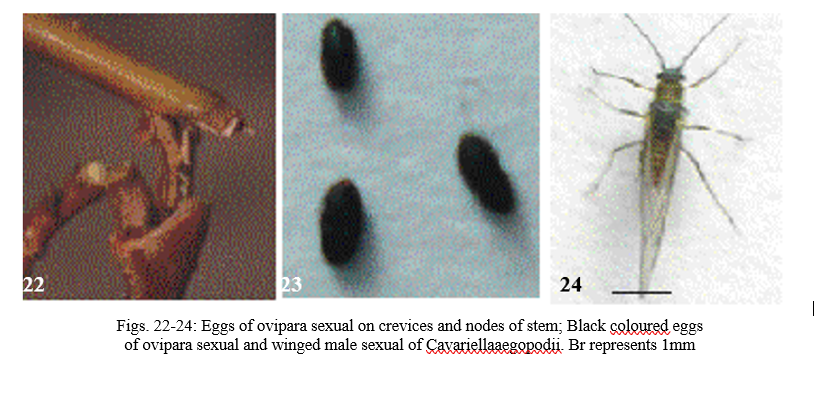
Ovipara sexuals occur in autumn season (September-October). The first instar (Fig. 17) is orangish brown in colour and is active right from birth. The second (Fig. 18) and third (Fig. 19) instars are orange brown in colour. The fourth instar (Fig. 20) has dark brown elongated body with swollen abdomen. Antennae are six segmented. Eyes are dark black in colour. The adult ovipara sexual (Fig. 21) is dark orange brown in colour and has elongated but swollen body. Adult ovipara sexual after mating usually searches for a suitable place to lay eggs in stem crevices, nodes and leaves (Fig. 22). While laying eggs, it raises posterior part of abdomen. Adult ovipara stops feeding after laying the eggs. Freshly laid eggs are creamish green in colour. The colour changes to light brown, and then to dark brown and finally the eggs turn black (Fig. 23). Eggs are laid in clusters and hatch in next spring.

Male sexual
Males occur in autumn season (September-October). Males are winged and black in colour (Fig. 24). The external male genitalia are heavily pigmented. Males are smaller in size in comparison to winged parthenogenetic female and winged female sexuparae. Males generally have a pattern of body markings similar to that of parthenogenetic winged females and have many more secondary sensoria on the antennae.
LIFE HISTORY OF DIFFERENT MORPHS OF CAVARIELLA AEGOPODII
The life history of Cavariellaaegopodiiwas studied in spring (March-April), summer (May-June), autumn (September-October) and winter (December to Feburary) to ascertain the duration of various stages of different morphs.
Parthenogenetic wingless female
The detailed life history of parthenogenetic wingless female was studied in spring, summer, autumn and winter seasons. The duration of first instar lasts from 2.5 to 3.5 (mean value 3.1) days in spring, 2.5 to 3.5 (mean value 3.2) days in summer, 3.5 to 4.5 (mean value 4.0) days in autumn and 4.5 to 6.5 (mean value 5.9) days in the winter (Table 2). The duration of second instar ranges from 3 to 4 (mean value 3.5) days in spring, 2.5 to 3.5 (mean value 3.0) days in summer, 2.5 to 3.5 (mean value 3.0) days in autumn and from 6 to 7.5 (mean value 6.5) days in winter (Table 2). The third instar stage lasts from 1.5 to 2.5 (mean value 2.2) days in spring, 2 to 3.0 (mean value 2.5) days in summer, 2 to 3 (mean value 2.5) days in autumn and 8 to 12 (mean value 9.6) days in winter (Table 2). The duration of fourth instar stage ranges from 3 to 4 (mean value 3.5) days in spring, 3 to 3.5 (mean value 3.2) days in summer, 1.5 to 2.5 (mean value 2) days in autumn and from 12 to 16 (mean value 14) days in winter (Table 2). The total duration of nymphal period of parthenogenetic wingless female ranges from 11 to 13.5 (mean value 12.3) days in spring, 11 to 12.5 (mean value 11.8) days in summer, 10.5 to 12 (mean value 11.5) days in autumn and from 33.5 to 37 (mean value 35.4) days in winter (Tables 2). The period between the last moult and the beginning of the reproductive period ranges from 2 to 4 (mean value 3.1) days in spring, 0.5 to 1 (mean value 0.8) days in summer, 1 to 1.5 (mean value 1.2) days in autumn and from 1 to 2.5 (mean value 1.5) days in winter. The total pre-reproductive period ranges from 13.5 to 17.5 (mean value 15.4) days in spring, 11.5 to 13 (mean value 12.6) days in summer, 11.5 to 13.5 (mean value 12.7) days in autumn and from 36 to 38.5 (mean value 36.9) days in winter (Table 2; Fig. 25).
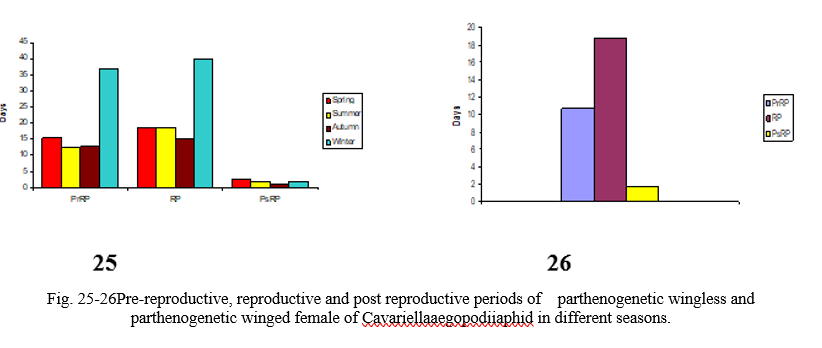
The duration of reproductive period in different seasons ranges from 11 to 26 (mean value 18.8) days in spring, 14 to 27 (mean value 18.8) days in summer, 11 to 20 (mean value 15) days in autumn and from 30 to 56 (mean value 40.1) days in winter (Table 2; Fig. 25). The progeny produced by parthenogenetic female varies in different seasons. In spring, a female lays 30 to 65 young ones in the whole life period (Table 2). In summer and autumn seasons, the progeny produced ranges from 35 to 75 nymphs and 30 to 45 nymphs respectively (Table 2). However, in winter a female produces only 19 to 30 nymphs in the whole life period (Table 2). Parthenogenetic wingless female takes about 20 to 45 minutes to lay the young one. It raises its posterior end and slightly jerks it at the time of giving birth to the young one. The maximum number of individuals produced by a parthenogenetic wingless female in a day is 5 in spring, summer and autumn seasons while it is 3 in winter season (Table 2).
Table 1: Record of temperature and relative humidity in the laboratory research chambers where biological studies were conducted from 2016 to 2019.
Year | 2016 | 2017 | 2018 | 2019 | ||||||||||||
Month | Mean Max Temp | Mean Min Temp | %RH | Mean Max Temp | Mean Min Temp | %RH | Mean Max Temp | Mean Min Temp | %RH | Mean Max Temp | Mean Min Temp | %RH | ||||
Max | Min | Max | Min | Max | Min | Max | Min | |||||||||
January | 12.1 | 3.5 | 76.16 | 64.96 | 13.8 | 4.8 | 56.2 | 43.5 | 12.2 | 3.5 | 71.9 | 60.5 | 11.7 | 2.4 | 89.4 | 79.4 |
February | 13.9 | 4.2 | 69.6 | 52.9 | 13.03 | 4.2 | 78.4 | 63.3 | 15.4 | 6.3 | 60.4 | 44.6 | 11.8 | 3.1 | 81.2 | 69.5 |
March | 18.2 | 8.9 | 61.6 | 47.5 | 17.3 | 7.5 | 62.7 | 49.1 | 22.0 | 11.3 | 47.3 | 41.3 | 17.9 | 7.3 | 77.1 | 64.9 |
April | 23.0 | 13.2 | 60.5 | 54.6 | 23.3 | 12.3 | 49.4 | 40.8 | 24.1 | 13.8 | 53.4 | 45.2 | 22.0 | 11.7 | 61.9 | 56.5 |
May | 26.6 | 17.2 | 61.5 | 54.7 | 25.6 | 15.0 | 44.8 | 36.5 | 26.6 | 16.5 | 54.5 | 45.7 | 24.6 | 14.3 | 52.2 | 46.0 |
June | 25.8 | 16.8 | 71.5 | 63.2 | 27.3 | 17.03 | 67.2 | 54.9 | 24.6 | 15.3 | 82.7 | 76.5 | 27.7 | 17.2 | 64.2 | 55.9 |
July | 24.8 | 17.1 | 83.8 | 78.2 | 23.5 | 16.7 | 94.1 | 89.5 | 24.4 | 16.5 | 89.2 | 80.2 | 22.9 | 16.3 | 96.1 | 90.3 |
August | 22.6 | 16.5 | 89.6 | 86.4 | 27.3 | 16.4 | 92.3 | 86.5 | 22.9 | 16.0 | 98.1 | 89.4 | 24.0 | 16.2 | 93.2 | 88.5 |
September | 22.4 | 13.6 | 82.7 | 74.1 | 23.1 | 14.9 | 86.5 | 78.1 | 24.3 | 15.3 | 79.9 | 68.2 | 23.6 | 15.2 | 86.6 | 80.2 |
October | 21.5 | 11.9 | 62.3 | 51.3 | 21.4 | 11.5 | 65.6 | 55.2 | 19.6 | 10.0 | 77.1 | 65.2 | 21.5 | 11.7 | 69.4 | 57.9 |
November | 18.2 | 9.5 | 59.0 | 42.0 | 17.8 | 7.8 | 71.6 | 58.2 | 17.3 | 8.5 | 70.7 | 61.2 | 17.6 | 8.1 | 60.2 | 52.4 |
December | 15.1 | 6.0 | 62.6 | 48.1 | 14.5 | 5.3 | 68.5 | 55.0 | 16.4 | 6.9 | 78.5 | 63.2 | 15.2 | 3.1 | 63.6 | 51.5 |
The post-reproductive period lasts for 2 to 3.5 (mean value 2.5) days in spring, 1.5 to 2 (mean value 1.8) days in summer, 1 to 2 (mean value 1.3) days in autumn and from 2 to 4 days with an average value of 1.8 days in winter (Table 2; Fig. 25). During this period, the parthenogenetic wingless female generally remains at a place and does not move around. It stops feeding and gradually dies. The duration of the total life period of parthenogenetic wingless female ranges from 29.5 to 43 (mean value 36.7) days in spring, 28.5 to 41.5 (mean value 33.2) days in summer, 25 to 34 (mean value 29) days in autumn and from 69 to 94 days with an average value of 78.8 days in winter (Table 2).
Parthenogenetic winged female
The life history of parthenogenetic winged female was studied during summer (May-June). The duration of first, second, third and fourth instar ranges from 2 to 3 (mean value 2.7) days, 2 to 3 (mean value 2.5) days, 2 to 2.5 (mean value 2.1) days and from 2.5 to 3.5 (mean value 2.7) days respectively (Table 2). The total duration of nymphal period ranged from 9 to 11 (mean value 9.9) days. The period between the last moult and the beginning of the reproductive period ranges from 0.5 to 1 (mean value 0.8) day. The total pre-reproductive period ranges from 9.5 to 11.5 (mean value 10.7) days (Table 2; Fig. 26). The duration of reproductive period ranges from 15 to 25 (mean value 18.8) days (Table 2; Fig. 26). The post-reproductive period lasts from 1 to 2.5 (mean value 1.7) days (Table 2; Fig. 26). The total progeny produced by a female in the whole life period ranges from 35 to 50 (mean value 41.3) days. Maximum number of nymphs laid by a female in a day ranges from 3 to 5 (mean value 3.9) days. The total life period ranges from 27 to 38 (mean value 31.2) days (Table 2).
Winged female sexupara
At the end of July month, the alate female sexuparae start appearing in parthenogenetc wingless colonies. Winged female sexuparae also undergo four moults. In the first and second instar nymphs of female sexuparae, wing pads are not visible. The wing pads become visible in third instar. The number of alate sexuparae starts increasing in the month of August and the maximum sexuparae population appears in September-October, both in nature as well as on plants maintained under laboratory conditions. Thereafter, their number starts decreasing in the month of November and continues till Feburary. The life history of winged female sexupara was studied in autumn season (September-October). The first and second instar stages last for 2 to 5 (mean value 3.6) days and 1 to 4 (mean value 2.7) days respectively. The duration of third and fourth instar stages ranges from 2 to 4 (mean value 2.4) days and 3 to 5 (mean value 3.8) days respectively (Table 2). The total nymphal period ranges from 10 to 14 (mean value 12. 4) days. The adult individuals live for 8 to 14 (mean value 11.1) days (Table 2). The winged sexuparae are parthenogenetic, viviparous and these give birth to apterous oviparae. The progeny produced by winged sexuparae range from 4 to 8 female sexuals i.e. oviparae. The total life period of sexuparae ranges from 20 to 27.5 days with an average value of 23.5 days (Table 2).
Table 2: Comparative chart showing the duration of different life history stages of different morphs of the Cavariellaaegopodii under laboratory conditions.
| Sr No. |
Character |
Parthenogenesis wingless female
| Parthenogenetwingedfwmale
Summer
| Winged female sexupara
Sept-Oct |
Sexuals
Ovipara Males
| ||||
| Spring Summer Autumn Winter | |||||||||
| 1 |
Duration of nymphal period (in days) |
I 2.5- 3.5
II 3 - 4
III 1.5-2.5
IV 3 - 4 | 2.5 – 3.5
2.5 – 3.5
2 - 3
3 – 3.5 | 3.5 - 45
2.5-3.5
2-3
1.5 – 2.5 | 4.5 – 6.5
6 – 7.5
8 - 12
12 - 16 | 2.0-3.0
2.0-3.0
2.0-2.5
2.5-3.5 | 2 – 5
1 – 4
2 - 4
3 - 5 | 2 – 2.5
2 - 3
2 – 2.5
2 - 3 | 2 – 3
2 – 3.5
3 - 4
2 – 3.5 |
2
|
Total nymphal period (in days) |
11 – 13.5 |
11 – 12.5 |
10.5 - 12 |
33.5 - 37 |
9.0-11 |
10 - 14 |
8 - 10 |
9 – 12.5 |
3 |
Period between the last moult and beginning of reproductive period (in days) |
2 - 4 |
0.5 - 1 |
1.0 – 1.5 |
1.0 – 2.5 |
0.5-1.0 | |||
4
|
Total pre-reproductive period (in days)
|
13.5 – 17.5 |
11.5 - 13 |
11.5 –13.5 |
36 – 38.5 |
9.5- 11.5
|
8 – 14*
|
2 – 8* |
4 -9* |
5 |
Reroductive period (in days)
|
11 - 26 |
14 - 27 |
11 - 20 |
30 - 56 | 15-25 | |||
6
|
Post – reproductive period (in days) |
2.0 –3.5 |
1.5 - 2 |
1 – 2.5 |
2.0 - 4 | 1.0-2.5 | |||
7
|
Total life period in days) |
29.5 – 43 |
28.5-41.5 |
25 - 34 |
69-94 |
27-38
|
20-27.5
|
11 - 17 |
13 – 21.5 |
| 8 |
Progeny laid by a female in the whole life period
|
30 - 65 |
35 – 75 |
30-45 |
19 - 30 |
35- 50 |
4 - 8 | ||
Ovipara sexual
The life history of ovipara sexual was studied in autumn season. The duration of the first, second, third and fourth nymphal instars varies from 2 to 2.5 (mean value 2.2) days, 2 to 3 (mean value 2.3) days, 2 to 2.5 (mean value 2.2) days and 2 to 3 (mean value 2.6) days respectively (Table 2). The total nymphal period ranges from 8 to 10 (mean value 9.2) days. The total life period ranges from 11 to 17 days with an average value of 14.3 days . The adults live for 2 to 8 days (Table 2).
Ovipara lays 3 to 8 eggs. The freshly laid eggs are creamish green in colour but later on, their colour changes to dark brown and then to black. The time interval between subsequent laying of eggs may range from 3 to 6 hours. Abdomen gets shrunk after laying of eggs.
Male sexual
Males of Cavariellaaegopodii are more active in comparison to ovipara sexuals. The life history of male sexual was studied in autumn season (September-October). The duration of first, second, third and fourth nymphal instars ranges from 2 to 3 (mean value 2.5) days, 2 to 3.5 (mean value 2.4) days, 3 to 4 (mean value 3.3) days and 2 to 3.5 (mean value 2.8) days respectively (Table 2). The total nymphal period ranges from 9 to 12.5 (mean value 10.9) days. The adults live for 4 to 9 days (Table 2). Males move about very actively to search for an appropriate mate. The total life period of male ranges from 13 to 21.5 days with an average value of 18.7 days (Table 2).
Cavariella del Guercio is reported to be a northern hemisphere genus of about 30 species, half of these species occur in Asia (Blackman and Eastop, 1994). Cavariellaaegopodii is widespread throughout the temperate and warm temperate regions and is reported to be heteroecious and holocyclic in most parts of the world, but often anholocyclic on Umbelliferae in warmer climates (Muller and Scholl, 1958). In India, Cavariellaaegopodii has been reported from Northwestern and Northeastern Himalayas (Raychaudhuri, 1980). Its detailed biology and cytogenetics have not been studied from India. The behavior and biology of this insect pest differs considerably in different regions due to different environmental conditions and present investigations revealed many interesting findings of its biology and cytogenetics in this region.
In Britain, primary host is Salix spp. and secondary hosts are cultivated species of Umbelliferae. Various morphs of Cavariellaaegoodii reported on these host plants are unwinged females, winged emigrant females, winged females, remigrant females, egg laying females and males (Dunn, 1965; Dunn and Kirkley, 1966 and Kundu and Dixon, 1994).
Present studies indicate that in this region, Cavariellaaegopodii has adapted to complete its life cycle on the primary host i.e. Salix spp. only. Life history studies of Cavariellaaegopodii revealed that in this region, this aphid species has different morphs in its life cycle. These are parthenogenetic wingless female, parthenogenetic winged female, winged female sexuparae, oviparae and male sexuals (Fig. 1). All the morphs undergo four regular moults resulting in five instar strages. The parthenogenetic wingless females occur throughout the year. Duration of nymphal instars varied in different seasons (Table 2) which is longer in winter season as compared to other (spring, summer and autumn) seasons. The total nymphal period (Table 2) of parthenogenetic wingless female is longer in winter (33.5 to 37 days) as compared to spring (11 to 13.5 days), summer (11 to 12.5 days) and autumn (10.5 to 12 days) seasons (Table 2). The period between the last moult and the beginning of reproductive period is more in spring (2 to 4 days) in comparison to summer (0.5 to 1.5 days), autumn (1 to 1.5 days) and winter (1.0 to 2.5 days) seasons (Table 2). The total pre-reproductive (36 to 38.5 days) and reproductive periods (30 to 56 days) are longer in winter season (Table 2; Fig. 25). Maximum progeny was laid in summer (35 to 75) and minimum in winter season (19 to 30) (Table 2). The longer pre-reproductive and reproductive periods and low fecundity during winter season (Table 2; Fig. 25) may be because of the effect of temperature and nutrition on the development of these aphids. The favourable conditions generally result in rapid development and production of more offspring. In spring and summer seasons, favourable temperature and nutrition result in rapid development of aphids and their high fecundity.
The life history of parthenogenetic winged female was studied during summer (May-June) season. Observations revealed that large colonies of aphids result in production of parthenogenetic winged females amongst the parthenogenetic wingless colonies. The parthenogenetic winged females were found to disperse to new plants and this resulted in new infestation. The total nymphal period of parthenogenetic wingless female (11 to 12.5 days) is more than parthenogenetic winged female (9 to 11 days) in summer (Table 2) and the period between the last moult and the beginning of reproductive period for both parthenogenetic wingless and parthenogenetic winged female is almost the same (0.5 to 1 day) in summer (Table 2). The progeny laid by parthenogenetic wingless female is more (35 to 75 nymphs) in comparison to the progeny laid by parthenogenetic winged female (35 to 50 nymphs) in summer (Table 2). The total pre-reproductive and reproductive periods for parthenogenetic winged female are 9.5 to 11.5 and 15 to 25 days respectively (Table 2; Fig. 26), whereas, these are 11.5 to 13 and 14 to 27 days respectively (Table 2; Fig. 25) for parthenogenetic wingless female during summer. The post-reproductive period for both parthenogenetic wingless female and parthenogenetic winged female is almost same (1.5 to 2 days) during summer season (Table 2). Similarly, the total life period for both parthenogenetic wingless female (33.2 days) and parthenogenetic winged female (31.2 days) is almost the same during summer season (Table 2).
At the end of July, the alate female sexuparae start appearing in apterous parthenogenetc colonies. The nymphal period of the alate female sexuparae (10 to 14 days) is longer (Table 2) than that of apterous parthenogenetic female (10.5 to 12 days) in autumn season (Table 2). The total adult life period for female sexupare varies from 8 to 14 days (Table 2). The fecundity rate of alate female sexuparae (4 to 8 female sexuals) is much lower as compared to parthenogenetic wingless female (31 to 45 nymphs) in the autumn season (Table 2).
Salix spp. is utilized as primary host and Umbelliferae and related Araliaceae as secondary host by all host alternating species of aphid Cavariella (Hille Ris Lambers, 1950, 1979). However, in this region, Cavariellaaegopodii has fully adapted itself on its primary host Salix babylonica. This may be due to the fact that some primary host plants continue to grow throughout the summer and are favourable from nutrition point of view supporting the earlier findings of Dixon and Glen (1971).
India has diverse climatic conditions and aphids generally reproduce parthenogenetically. Sexual morphs have been recorded in very few aphid species. In the present investigations, sexual morphs of Cavariellaaegopodii were recorded in natural populations. The life history of sexual morphs i.e. ovipara and male sexual was studied in autumn season (Table 2). In male sexuals, the total nymphal period ranges from 9 to 12.5 (mean value 10.9) days (Table 2). The adult males live for 4 to 9 days (Table 2). The total life period of male ranges from 13 to 21.5 days with an average value of 18.7 days (Table 2). However, the total nymphal period of ovipara sexual ranges from 8 to 10 (mean value 9.2) days (Table 2). The total life period of ovipara ranges from 11 to 17 days with an average value of 14.3 days (Table 2). An oviara lays 3 to 8 eggs which are black in colour (Table 2).
In the present study, maximum number of oviparous females and males was observed during autumn season. It is interesting to note that sexuales were collected from September to October when the day length is relatively longer and temperature is higher than during winter. The occurrence of sexuales during colder part of the year (winter), when the day length is rather short and temperature low, is in conformity with the observations of earlier workers on other aphid species (Macrovitch, 1924; Kenten, 1955; Lees, 1959, 1960, 1963; Lamb and Pointing, 1972; Sharma et al., 1975; Matsuka and Mittler, 1979; Brodel and Schaefers, 1980), although these have not been considered as the only determining factor for the production of sexuales by other workers (Bodenheimer and Swirski, 1957). The mechanism of photoperiodic response has not been studied in Cavariellaaegopodii. In Cavariellaaegopodii, sexual morphs occurred in autumn season confirming the early reports by Bonnemaison (1951), that the short-day length of autumn season is conducive for sexual morph production.
There is no clear evidence that any environmental factors except photoperiod and temperature have a direct influence on this aphid life cycle. Oviparous females are apterous and orange brown in colour while males are winged and black in colour. The total nymphal period (8 to 10 days) of oviparous female is shorter than that of males (9 to 12.5 days) (Table 2). Similarly, the total life period of oviparous female is shorter than that of males (Table 2). Production of sexuals is attributed to some intrinsic timing mechanism (Bonnemaison, 1951) or interval timer as reported by Lees (1960). Kundu and Dixon (1993) reported that the remigrant females of Cavariellaaegopodii on willow survive for 30 days. Males start feeding on willows on first or second day after their adult moult, however, it is reported that males on willow probably start feeding 5 days after the adult moult. Dunn (1965) reported the presence of eggs on Salix alba and Salix fragilis but no eggs were recorded on Salix babylonica by him. However, in this region, eggs were found on Salix babylonica host plants.
Blackman, R.L. 1980b. Chromosomes and parthenogenesis in aphids. Symposia R. Ent. Soc. Lond. 10: 133-148.
Blackman, R.L. and Eastop, V.F. 1994. Aphids on the world's trees. CAB International, Wallingford, U.K. pp 466
Bodenheimer, F.S. and Swirski, E. 1957. The Aphidoidea of the Middle East. The Weizmann Science Press, Jerusalem
Bonnemaison, L. 1951. Contribution al étude des facteurs provoquant l apparition des formes ailees et sexuees chez les Aphidinae. Ann. Epiphyt. (Paris), 2: 1-380
Brodel, C.F. and Schaefers, G.A. 1980. The influence of temperature on the production of sexuals by Aphis rubicola under short–day conditions. Entomol. Exp. Appl. 27: 27-132.
Dixon, A.F.G. and Glen, D.M. 1971. Morph determination in the bird cherry-oat aphid Rhopalosiphumpadi. Annals of Applied Biology, 68: 11–21
Dunn, J.A. 1965. Studies on the aphid Cavariellaaegopodii Scop. I. On willow and carrot. Ann. Appl. Biol. 56: 429-38
Dunn, J.A. and Kirkley, J. 1966. Studies on the aphid Cavariellaaegopodii Scop. II. On secondary host other than carrot. Ann. Appl. Biol. 58: 213-217
Hille Ris Lambers, D. 1950. Host plants and aphid classification. Proceedings. 8th International. Congress of Entomology, Stockholm, Sweden, 1948. pp141–148.
Hille Ris Lambers, D. 1979. Aphids as botanists? SymbolaeBotanicaeUppsaliensis, 22: 114-119.
Holdenrieder, O.; Rudow, A. and Aas, G. 1999. The genus Salix: An introduction to the diversity, biology and ecology of willows. Schweizerische-Zeitschrift fur Forstwesen. 150: 405-411.
Kennedy, J.S.; Day, M.F. and Eastop, V.F 1962. A conspectus of aphids as vectors of plant viruses. Lond. Comm. Inst. Entomol. pp1-144
Kenten, J. 1955. The effect of the photoperiod and temperature on reproduction in Acyrthosiphonpisum Harris and on the forms produced. Bull. Entomol. Res. 46: 599-624.
Kundu, R. and Dixon, A.F.G. 1993. Do host alternating aphids know which plant they are on? Ecological Entomology, 18: 61-66
Kundu, R and Dixon, A.F.G. 1994. Feeding on their primary host by the return migrants of the host- alternating aphid, Cavariella aegopodii. Ecological Entomology, 19: 83-85.
Kurzovkina, Y. A. and Quigley, M. F. 2005. Willows beyond wetlands: uses of Salix L. species for environmental projects. Water, Air and Soil Pollution. 162: 183-204.
Lamb, R.J. and Pointing, P.J. 1972. Sexual morph determination in the aphid, Acyrthosiphonpisum. Journal of Insect Physiology, 18: 2029-2042
Lees, A. D. 1959. The role of photoperiod and temperature in the determination of parthenogenetic and sexual forms in the aphid, Megouraviciae Buckton. I. The influence of these factors on apterous virginoparae and their progeny. J. Insect Physiol. 3: 92-177.
Lees, A. D. 1960. The role of photoperiod and temperature in the determination of parthenogenetic and sexual forms in the aphid, Megouraviciae Buckton. II. The operation of the “interval timer” in young clones. J. Insect Physiol. 4: 154-175.
Lees, A.D. 1963. The role of photoperiod and temperature in the determination of parthenogenetic and sexual forms in the aphid, Megouraviciae Buckton. III. Further properties of the maternal switching mechanism in apterous aphids. J. Insect Physiol. 9: 153-154.
Macrovitch, S. 1924. The migration of the Aphididae and the appearance of sexual morphs as affected by the relative length of daily light exposure. J. Agric. Res. 27: 513-522.
Matsuka, M. and Mittler, T.E. 1979. Production of males and gynoparae by apterous viviparae of Myzuspersicae continuously exposed to different scotoperiods. J. Insect Physiol. 25: 587-593.
Muller, F.P. and Scholl, S.E. 1958. Some notes on the aphid fauna of South Africa. J. Entomol. Soc. S. Afr. 21: 3.
Raychaudhuri, D.N.1980. Aphids of North-East India and Bhutan. The Zoological Society, Calcutta.
Sharma, M.L.; Larrivee, J.M. and Theriault, L.M. 1975. Sequences de descendence des formssexuees chez le puceron du pois Acyrthosiphonpisum(Homoprtera : Aphididae) en relation avec la duree des photoperiodes. Canadian Entomologist, 107: 1063-1067.
