
+91 6002993949
submission@iarconsortium.org
Open Access
ISSN (Print) : 2708-5090
ISSN (Online) : 2708-5104
This is a review paper assessing the ecological resilience of the Luangwa (Zambia) hippopotamus (Hippopotamus amphibius) with respect to (i) Population Size, Structure and Distribution, (ii) Primary Production and Grazing Capacity, (iii) Mortality and (iv) Population distribution in relation to non-food resources and security. The information analyzed in this report was based on the data collected since 1976 using total river bank counts. In this technique a pair of binoculars and GPS locations are used to record schools and to classify them in age groups. Records from culled carcasses are also used to consolidate sex and age classes’ determination. Data on body condition is based on Kidney Fat Index (KFI), while mortality records kept at Chinzombo Research stations covers deaths from hunting, culling, control, disease and senescence as unknown. Data on pasture is based on the clip-weigh - dry - re-weigh method tied to the Becovol Model and used to obtain above ground biomass, while the influence of the river geomorphology in creating a favorable habitat is based on satellite imagery and ground. As reported by several earlier studies, the Luangwa hippo population is resilient. Supported by the good quality habitat with numerous geomorphologic features which among other things enhances calf survival while habitat loss is insignificant because a significant proportion of the river is located in National Parks and Game Management Areas. The proposal to uplift the Appendix listing of the hippo population from Appendix II to Appendix I of CITES is not valid for Zambia. International trade of hippo specimens is not a significant factor affecting the hippo population in Zambia. Attention should be placed on habitat quality, anthropogenic influences such as encroachment of humans on hippo habitat and perhaps water availability in drought years and the effects of Climate change.
The common hippopotamus population of the Luangwa valley has been resilient for a period exceeding a century now. It has since the mod 1970s recorded high densities of up to 48 hippopotami per kilometer of the river, usually exhibiting a rise and fall which has persisted since the 1970s. Culling and anthrax, such as the outbreak of 1987-1988, reduced the population to below 5,000 but the population rose again to pre anthrax levels by 1989 making the population resilient to environmental resistance. Poaching and natural mortality including predation and senescence have had no significant impact in reducing the population. Every time there is a decline, it is followed by increases. This resilience has prompted management to conduct culling as one way of reducing the population while providing meet and trophies as an additional economic incentive. During the period 2005 – 2008 [1], a culling was carried out after a protracted engamenet of local and international stakeholders from the two schools of thought. The anti-consumptive utilization and the consumptive utiulisation. Each school with convincing data. The data from this culling was collated and stored. In 2016 another culling was attempted but this one dismally failed due to local and international resistance from animal rights groups and anti-consumptive utilization schools of thought. International media, including selected foreign missions using their widespread networks forced govenemnent the plan of ciliing. The population howver continued to rise. The question that reapeatedly arises is, what special biophysical factors are responsible for the resilience of this population?
To answer this question, a through literature review of existing data covering the biology, ecology and physical factors were reviewed.
Environmental factors and anthropogenic issues such as trophy hunting and international trade and culling human–hippopotamus conflicts, disease outbreaks, poaching, primary production and rainfall security and many others were also reviewed. We also sparingly compared with other populations including those of the pygmy hippo.
Below we start reviewing the biological factors of the species and end with social factors.
General Description of Hippopotamus
Common hippopotamus (Hippopotamus amphibius Linnaeus, 1758), (Figure 1) commonly called hippo is an English colloquial name derived from Greek, which means “water or river horse”. This is however, is a misnomer as hippos are not related to horses. In fact, the hippopotamus was previously thought to be more closely related to pigs than to horses [2-4]. This classification was based on molar patterns, but in 1985 evidence from blood proteins, followed by molecular systematics and more recently fossil evidence, has shown that the closest living relatives are cetaceans – whales and porpoises [5-7].
This classification, however, still presents challenges to taxonomists as both cetaceans and hippopotami are early offshoots of artiodactyls evolution. The whale and hippopotamus lineages split very soon after they separated from the rest of the artiodactyls. In this regard, the hippopotami classification still remains a subject of controversy [7].
The species is characterized by large size, barrel-shaped body, smooth and naked skin and short stout legs (Figure 1). The head is broad and heavy with the eyes and nostrils ‘mounted’ on top [4]. The mouth is wide and large; the jaws are deeply cleft and armed with tusk like canines and incisor teeth which are a major source of ivory and an area of contention in international trade regulated under CITES and perhaps the major reason for the proposed upscaling to Appoendix I of CITES. In proportion to the head, the ears are small and upstanding (Figure 1). The eyes, nostrils and ears are located on the upper part of the head as adaptation for an amphibious life, soi that the individual can see and breathe while almost the entire body is submerged in water for cooling.
The dental formula for an adult is:
I 2/2, C 1/1, P 3/3-4/4, M 3/3 = 36 – 40
While the cheek teeth are adapted to grinding, the positioning of the canines and incisors renders them useless in feeding (Figure 2).
Canines and incisor teeth do not play any role in grazing and the cropping of grass is done by hard edges of the lips while canines and incisor teeth function as weapons of defense and display, the popular one being the yawn display.
Body length is 320-420 cm for males and 280-370 cm for females. Height is 140 – 165 cm for males and 130 - 145 cm for females. Weight is 1,500-3,200 kg for males and 1,350 - 2,500 kg for females [2,4,8]. Larger specimens of up to 5,000 kg measuring five metres long have been recorded [5]. With such body measurements hippos are said to be approximately the same size as the White Rhinoceros (Ceratotherium simum) and experts are split on which is the next largest land animal after the elephant (Loxodonta africana). Male hippopotami in particular appear to continue growing throughout their lives, whereas females reach a maximum weight at about the age of 25 years [2].
Taxonomy
The area of taxonomy is another potyential locus of dispute and particularly when matters of CITES are in focus. However, authorities recognize seven sub species five in the genus Hippopotamus
H.a. amphibius (considered to be the nominate race by taxonomists)- East Africa and west to the Gambia
H.a. tschadensis - Chad and Nigeria
H.a. kiboko - Kenya and Somalia
H.a. constrictus- Angola and Namibia
H.a. capensis- Zambia and southwards to South Africa and two or three in the genus Hexaprotodon
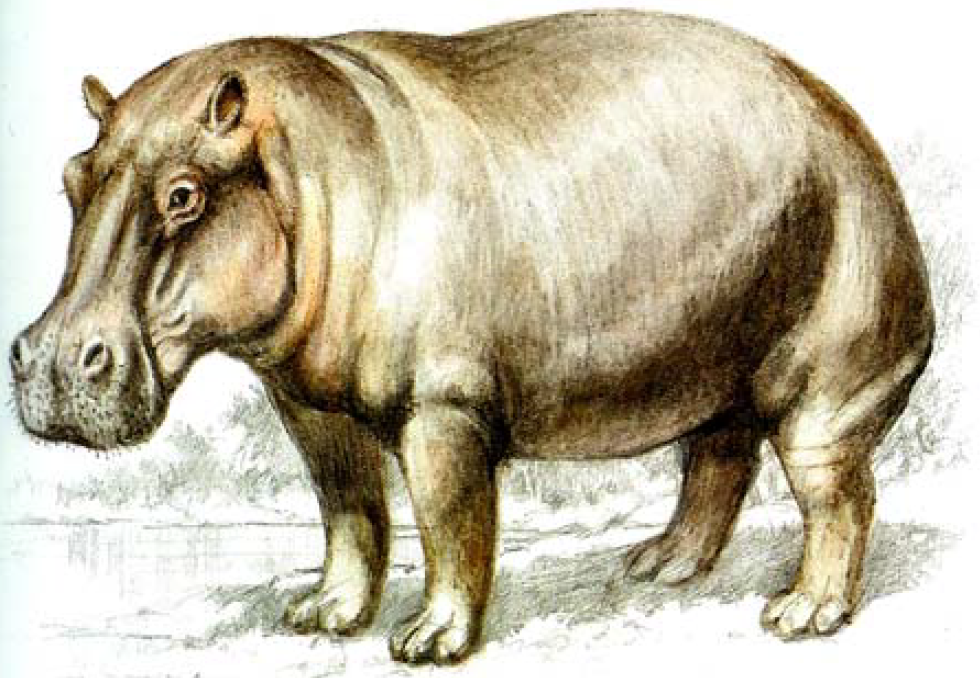
Figure 1: Common Hippopotamus
Source: [4]
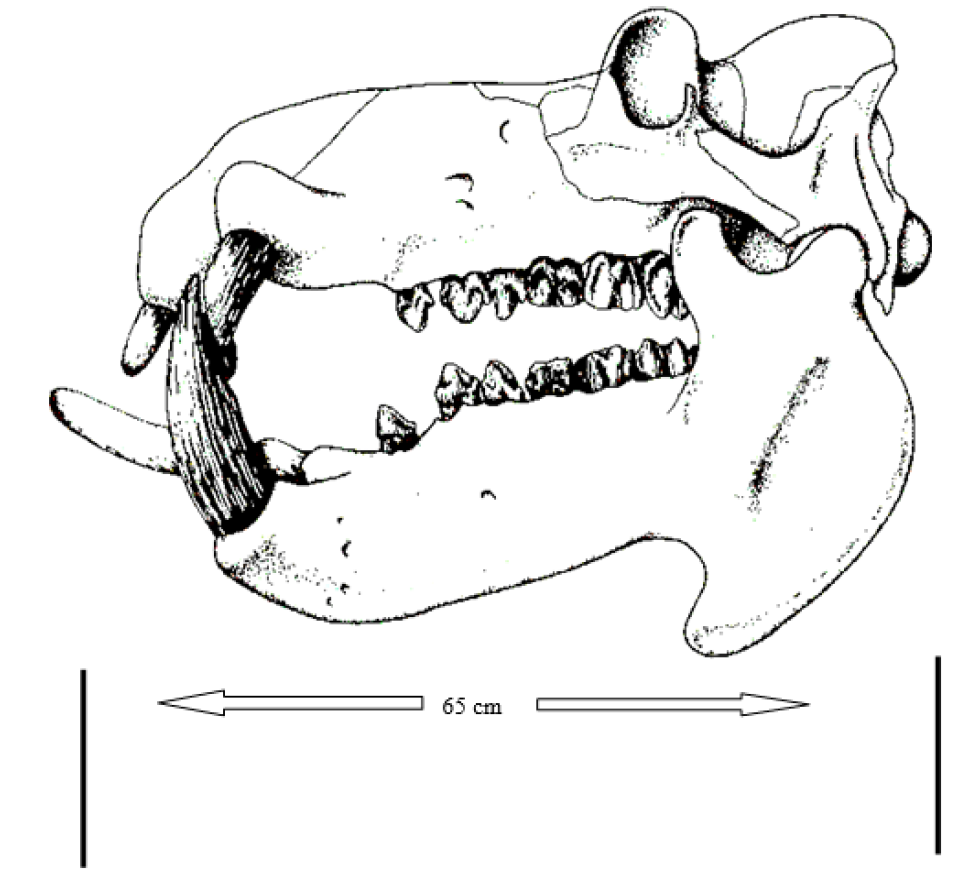
Figure 2: Skull of Common Hippopotamus Harvested from the Luangwa River Culling Scheme Showing Large Canines and Cheek Teeth (Total length of Skull was 65 cm), Luangwa Valley, Zambia
Which are, Hexaprotodon liberiensis that, is smaller than common hippopotamus, weighing about 270 kg and the shoulder height being 80 cm. Its distribution is restricted to West Africa. Unlike the common hippopotamus it has proportionally smaller, narrower head with orbits not raised above the skull roof. It has 38 teeth, compared with 42-44 in the common hippopotamus [9] (Figure 3).
The second is the nominate race, H.l. liberiensis which occurs in Guinea, Sierra Leone, Liberia and the Ivory Coast and H.l. heslopi which according to Corbert [10], occurs from Niger Delta east to the vicinity of the Cross River in Nigeria. It is distinguished from the nominate subspecies by smaller skull proportions [11,12]. The Luangwa hippo which is the population of interest is therefore H.a. capensis.
The classification of common and pygmy hippopotami into subspecies still remains a subject of controversy until comprehensive molecular analysis is done on all subspecies. Above species level however, molecular data have supported a sister group relationship between hippopotamids and cetaceans [7].
Ecology and Behavior
Hippopotamus as its common name suggests, is an amphibious animal which spends most of the day time in water and mostly emerging at night to graze. Water is used as day time retreat and does not usually eat aquatic vegetation. It is assumed that the habit of feeding at night helps the species to avoid the direct day time rays of the sun which can dry and dehydrate its skin. It leaves water mainly soon after sunset and spends the night grazing on short grass lands for up to several kilometers (usually 2–5 km) from water. Grass on the grazing areas is kept short by activities of the hippopotamus and these areas are commonly known as hippopotamus grazing lawns (Figure 4).
Although open water is important for hippopotamus it can also survive in muddy wallows as long as there is access to permanent water to which it can return in the dry season. The essential role water plays are to keep the skin moist for it would crack if exposed to dry air for long periods [5]. The skin physiology is complex and not fully understood but is clearly adapted for amphibious existence. A curious feature is the red secretion from modified sweat glands, which is thought to have an antibiotic function and could be attributed to the species’ relative resistance to disease infestation [5].

Figure 3: Pygmy Hippo
Suitable water habitat for hippopotamus should be enough to accommodate social groups called schools which are infact unstable groups of females and bachelors formed on the basis social hierarchy, where territorial males monopolise a length of the shoreline of the river or lake but tolerate bachelors within the territory provided they behave submissively. It is for this reason that when assessing extent of hippopotamus habitat, as has been the case in the Luangwa River studies, it is always important to refer to bank length when calculating density and assessing population distribution. The meandering of the Lungwa River increases bank length creating abundant micro habtas for the species (Figure 5).
Males fight fiercely for territories often resulting into considerable body injury or death and non-territorial and un submissive males are usually kept outside territorial areas usually in seasonal wallows and other marginal areas. In most instances however, such fierce fights are avoided through threat displays exposing their huge canines by yawning [13]. It would appear therefore, that major fights occur within the school or when establishing a territory. At night, the schools break up and go on land to feed and there is little association except for females and their dependent young and the males cease to behave in a territorial manner [13]. The spreading of dung by tail wagging while defecating in water or on land is usually done by males and has no territorial function but has a signaling function for orientation. Vocalization is in form of complex bellows and grunts, which presumably also have a signaling function [5]. Sounds are made either on land or in the water and may be transmitted simultaneously through air and water Eltringham [13] and Saikawa, et al., [5], recorded this as being the only known case of amphibious calls in a mammal. In the Luangwa Valley, hippo schools of about 300 individuals (personal observation) are recorded, yet with occasional fights. The multiple channels and lagoons, perhaps may serve as dispersal areas for non dominat individuals and this may reduce levels of fights among dominat bulls. River meandering, formation of multiple channels and lagoons, could in this instance be consider to be a favouarable habitat attribute facilitating dispaersal of individuals and also as retreat areas for less dominat individuals. Such habiata attributes are rare in other water bodies such as the Kafue and Zambezi rivers, which though being beigger than the Luyangwa do not hold comoarable hippo popluations to the Laungwa river.

Figure 4: Hippopotamus ‘Grazing Lawn’ (Cynodon spp), Block B Nsefu Area Upper Study Area, Luangwa Valley, Zambia
Although hippopotamus usually grazes at night there are usually animals present in the water all night, as individuals do not leave water at the same time. Some leave early and return few hours later and then others leave also. When feeding, grass is passed to the back of the mouth where it is ground up by molars. McCullough, [2], noted that the amount of food ingested by hippo is small relative to its size but its resting habits by day reduce energy demands. It is for this reason that hippopotamus cannot do without open water. The stomach is a complex four-chambered structure with a ruminant type digestion although the animal does not chew the cud.
It can be said therefore, that the habitat requirements for hippopotamus in general, include a supply of permanent water large enough for the territorial males to spread out and adequate grazing open grassland within a few kilometers (2-5 km) of the daytime resting sites for which the Luangwa River offers in abundance (Figure 6a, b).
Habits
Hippopotamus live in social groups called schools (other names are pod, herd or bloat) which vary in size and composition; solitary animals are generally territorial bulls or females about to give birth. Generally, groups consist of females with calves, or bachelors in association.

Figure 5: River Meander, Minor Channel Which form River Cutoffs Study Block B. South Luangwa National Park, Zambia [40,41]
Figure 6a: Details of the Physiographic Map of the Luangwa Valley, Zambia Showing the Alluvial Belt Which Supports Grassland Plains as Hippo Pasture.
Size of schools vary from 10 -15 or up to 30. In the Luangwa valley, schools of up to 500 individuals have been recorded especially in the last quarter of the year (October–December) (personal exp.) when it is dry. Haltenorth and Diller [8] and McCalough [2], recorded groups of about 10-15 as being the commonest school sizes in most water bodies. Dorst and Dandelot [9] and Stuart and Stuart [14], recorded that during periods of drought large numbers congregate in few remaining pools of water and schools larger than 30 individuals would be found. Eltringham [13], noted that territorial fighting increased with increase in school size. It appears therefore, that overcrowding also disrupts the hierarchical system, resulting in higher levels of aggression. Under such conditions, breeding may be disrupted. This factor perhaps is part of the basis for culling to minimize aggression and promote birth rates. Males continue to grow throughout lifetime and the oldest and strongest males are usually the most dominant. For that reason, body size as dimorphic feature has been used in the Luangwa hippo studies to distinguish mature males from mature females.

Figure 6b: Hippopotamus Passture Comprising Short Grasslands with Stands of Undesirable Sporobolus spp and Andropogon spp, while Cynodon Dactylon was Grazed to Carpet Level. Block B in Nsefu of the Upper Study Area, South Luangwa National Park, Zambia

Figure 7: Hippo Foraging and Pathway from Flood Zone Through Tall Grassland to Grazing Lawns. Nsefu Area, South Luangwa National Park, Zambia
Grazing areas are not subdivided into territories and change with season and availability of forage [13]. To access grazing areas hippopotami, use established routes called hippopotamus pathways, where the ground eventually becomes bear (Figure 7). Where food is plentiful, they tend to remain in the vicinity of the resting pool but would move freely up to five or in some cases up to 10 km when food is scarce or during drought (personal exp.). With the meandering of the Luangwa River and the rich alluvial depoisits, foraging areas are always within in most instances within 5km of the river banks which makes the Luangwa River a very unique hippo habitat and perhaps also reposible in part for the super abundance of hippo not recorded anywhere among the range states.
Reproduction
Little is known about the hippopotamus’ general reproductive biology, dispersal phase of its life history and its population genetics. Eltringham [13], attributed the lack of information on hippopotamus reproductive biology to the semi aquatic nature and nocturnal habits of the species which makes it difficult to obtain representative samples. Puberty is attained at 7.5 years for females (females have been recorded to mature at four years) and 9.5 years (or less) for males [15,16]). The relatively short maturity period particulalrly females is one of the reasons for the quick recovery of popolations in areas where the habitat is suitable which applies to the Luangwa valley.
Mating in hippopotamus takes place in water or on land as earlier reported by Chomba et al. [17-19], after passing through a series of body contact courtship behaviours to entice the female. In pre-copulatory behaviour the male hippopotamus is not aggressive, but the female appears to walk away while the male pursues. Sometimes the female would walk considerable distances while the male follows behind. Shallow water appears to be the most preferred mating area (personal exp.); probably because the male is buoyant enough to mount a female with ease and can breathe normally. In four of our observations, three instances were in shallow water and only one on dry land signifying the importance of water in mating behaviour (personal observationbs). If in water the female remains submerged, except for brief intervals to breath, while the male’s head is not submerged.
Calving and Calf Survival in the Luangwa Valley
Gestation period is between 225-257 days as recorded by Asdell [20], Haltenorth and Diller [8] and McCalough [2] which is surprisingly much shorter when compared with other mammals of similar size; buffalo-330 days, black rhino -480 days, white rhino 540days and although normally one young is born every 11/2 – 2 years this a relatively short gestation period for a large mammal whose mean body weight exceeds 1,000 kg. Females have their first calves at four years old. Males reach puberty when they are about seven years as recorded by Skinner, Scorer and Millar [4]. When about to calve, the female leaves the group and finds a secluded site in shallow water to give birth to a single young. Twins have been recorded but are not common [8]. Birth takes place according to circumstance, either in shallow water or on land. If on land, the female makes a grass or reed bed in hollow of bank, away from the herd. Hind legs emerging first, which have a mass of about 50 kg as recorded by Haltenorth and Diller [8], Skinner and Smithers [4]. In the Luangwa Valley during the 1970-1971 culling programme a foetus of 50 kg was recorded by Sayer and Marshall [21]. Few minutes after birth it is capable of going into deep water but like adults, young hippopotamus are unable to float. The question is, was the Luangwa crafted by nature to hold hippo popuylaytions? So far, all hippo biological attributes seem to find space in the Laungwa valley, a factor not shared with other water bodies in the country and perhaps other regions of the distribution range.
Imprinting takes up to 14 days and is critical for establishing calf-mother relationship. At this moment, the female and young usually remains apart from any other members of the group. When the pair returns to join the group, the calf usually stays near the shore while the mother feeds in the vicinity. In the Luangwa Valley the calves are left on sand bars surrounded by water for security (personal experience) and this could be responsible for the high calf survival in the Luangwa valley. The meandering of the Luangwa River and the sand depositions which creates numerous sand isalands is reposnsible for the high calf survival (Figure 8a, b).
This also makes it easy during the river bank counts to record both adults and young ones.

Figure 8a: River Meanders and Sand Bars

Figure 8b: Sand Bars Providing Sanctuary for Calves’ Thereby Enhancing Calf Survival
Impact of Disease
Regarding susceptibility to disease, hippopotami are not very susceptible to diseases [5] and this is also true in the Luangwa Valley where anthrax out breaks have had no negative impact on the population density. Longevity is 40-45 years and up to 50 years in captivity [2,3]. With such inherent biological characteristics, in suitable habitats like the Luangwa valley in Zambia, their numbers can increase quickly in a relatively short period of time. In the Luangwa valley, outbreak of disease affecting hippo are occasional. The most common one is the anthrax. Overtime, the incidences of anthrax has been recorded after a drought [22-27]. During the years of drought, there is a limitation of pasture and the hippos graze close to ground levels and often pulling ut the grass roots of the grazed species of grass thereby exposing the anthrax spores which hide below ground, leading to an outbreak. Even after an outbreak and loss of individuals, the population bounces back failry quickly, owing to the short gestation period, reduced intercalving interval and abundance of pastrure. All previous records including Chomba [1] and Plowright et al., [27] suggest that the effect of disease on the Luangwa hippo population is insignificant. Whether this resilience to disease exhibited by the Luangwa hippo is common to other hippo range states is currently not ascertained and be subject of further research.
Feeding
Hippopotamus are selective nocturnal grazers, consuming 50 kg (40 - 60 kg) of grass or about 2.25% of their body weight daily [28]. They also occasionally scavenge meat from dead animals found near their range, though hippopotamus is not carnivorous in any realistic sense. In drought they can fast for some weeks [8], during which time body condition rating drops to poor. Types of grasses utilized by hippopotamus depend on grass availability, though Scotcher, Stewart and Breen [29] showed that in Ndumu Game Reserve, South Africa buffalo grass, Panicum maximum, Urochloa mosambicensis and couch grass Cynodon dactylon, occurred predominantly in their diet, in the other Hermathria altissima and Echinocloa pyramidalis. Around S’t Lucia, Ischaemum arcuatum was the most preferred species. Dorst and Dandelot [9], recorded Themeda, Panicum and Heteropogon and aquatic plants Pistia stratiotes. Fruits of Kigelia pinnata were also recorded. Hippopotamus are very close croppers and will graze on stands of grass such as couch grass, C. dactylon, until they are of lawn like appearance (Figure 9 and Figure 4). This is accomplished with the horny edges of the lips. The grass is plucked with an upward movement of the head. In the Luangwa Valley, pasture is rich with palatable species benefiting from the subsusrfce misture in the dry season keeping the hippos nourished, all year round except years of drought.
With the wide choice of forage, there is no doubt that the Luangwa valley is a favourable habitat for hippo. The alluvial belt provides a sutable and rich pasture for the hippo and is responsible for the superabundance of the species (Table 1).

Figure 9: The Horny Edges of the Lips Enables Hippo to Crop Grass to Near Carpet Level, Luangwa Valley

Figure 10: Gentle Slopes and Sand Bars Are Suitable Basking Areas for Hippopotamus Schools. Adults Bask on the Periphery to Protect Calves from Land Predators such as Lion. NSEFU Area, Study Block B, Luangwa Valley, Zambia
Habitat
Essential habitat requirement is sufficient open water in which they can submerge totally and cool their bodies, though they usually prefer shallow standing or slow moving water near sandy bars or banks on which they can bask in the sun (Figure 10). If undisturbed they can use these pools over a period of many years if food supplies remain available within their range of normal movement. Sometimes, seasonal flooding may cause them to move temporarily from their established resting pools if such flooding seriously alters pool condition; they usually return as the flood subsides. During high floods they find temporary resting places in lagoons or up the tributaries [5], of which there are plentiful in the Luangwa valley.
Area of Study
The data used in this review paper only covers the Luangwa valley in eastern Zambia which holds about 62% of the national hippo population (Figure 11). Located at (12o 48’S, 32o 03’E) to the Lusangazi-Luangwa confluence (13o 24’ S, 31o 33’ E).
Table 1: Herbaceous Species Grazed by Hippos in the Study Blocks A - H, Luangwa Valley, Zambia
| Species Name | Species sighting frequency | Signs of grazing | |
| Short grassland | Tall grassland | - | |
| Andropogon spp | - | 15 | - |
| Aristida adscensionis | - | 5 | Not grazed |
| Brachiaria spp | 5 | - | Grazed |
| Chloris spp | 10 | - | Grazed |
| Cenchrus ciliaris | 12 | 8 | Grazed |
| Cynodon dactylon | 25 | Grazed | |
| Cymbopogon spp | - | 33 | Not grazed |
| Cyperus rotundus | - | - | Grazed |
| Dactyloctenium | 5 | - | Grazed |
| Digitaria spp | 8 | - | Grazed |
| Echinocloa pyramidalis | 20 | - | Grazed |
| Diheteropogon | - | 6 | Not grazed |
| Commelina | - | Grazed | |
| Eragrostis superba | 5 | Grazed | |
| Heteropogon spp | - | 10 | Not grazed |
| Heteropogon contortus | - | 10 | Not grazed |
| Hyparrhenia | - | 9 | Not grazed |
| Hyperthelia | - | 4 | Not grazed |
| Panicum | 8 | - | Grazed |
| Phragmites australis | - | Not grazed | |
| Setaria | 5 | 7 | Not grazed |
| Sporobolus spp | 3 | - | Grazed |
| Themeda triandra | - | - | Not grazed |
| Total number of species | - | 12 | - |
| Number not grazed of total | 2 (14 %) | 11 (92 %) | - |
Source: [1,42], Blanks Represent Frequency of less than 5 %.
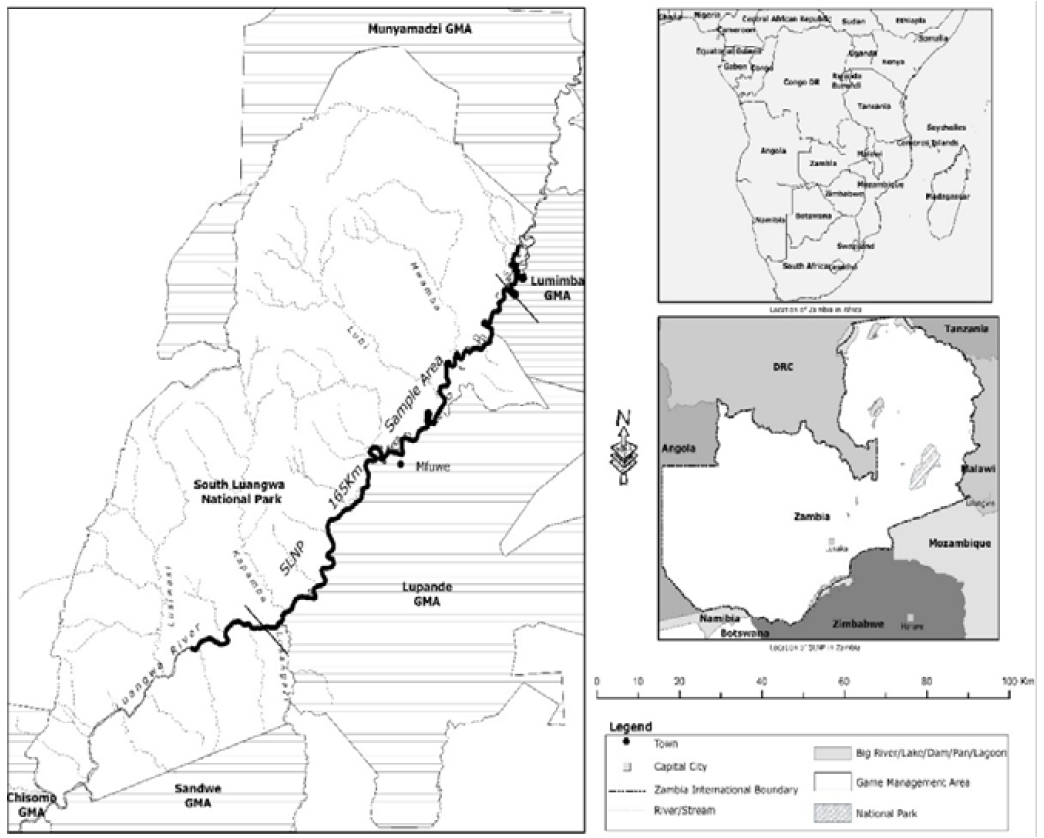
Figure 11: Location of Study Area Along Luangwa River and South Luangwa National
Distribution
All large rivers and laekes in Zambia have hippo population, but none exceeds the Luangwa River which has 62 % of the national hippopopulation [1,17-19]. Elsewhere, though not clear as the distribution is normally described on the basis on national political boundaries and not science it is fairly widespread through the numbers seem to me declining in the majority of range states.
The nominate race, Hippopotamus amphibius amphibius, is known to occur in southern Sudan, Ethiopia and northern Democratic Republic of Congo, Uganda extending west to Gambia and further south to Tanzania and Mozambique. Hippopotamus amphibius tchadensis occurs in Chad and Niger while Hippopotamus amphibius kiboko occur in Kenya and Somalia. Hippopotamus amphibius capensis is distributed from Zambia south to South Africa. Hippopotamus amphibius constrictus occurs in Angola, southern Democratic Republic of Congo and Namibia. As earlier said, the species’ boundaries are not well defined and significant overlaps may oiccur as supported by the study carried out by [30], in which an unrooted neighbour-joining tree based on net genetic distances showed relationships among the six hippopotamus populations sampled (Figure 12,13).

Figure 12: Schematic Map Showing 13 Sampling Localities of Hippopotamus in Eastern and Southern Africa. Sampling Sites, Population and Subspecies Groupings are: 1 Murchison Falls National Park (n = 28) of MNP Population; 2, Queen Elizabeth National Park (n = 25) of QEN Population; 3, Ugalla (n = 8); 4, Moyowosi (n = 1); 5, Luganzo (n = 1); Kigosi (n = 5); 7, Mlele (n = 1); 8, Niensi (n = 1); and 9, Selous (n = 1) of Tanzania (TZN) population and above Population Constitute H. a. Amphibius subspecies. 10, Masai Mara Game Reserve (n = 25) of MAM Population and 11, Naivasha National Park (n = 9) of NAV Population, Cover H. a. kiboko Subspecies. 12, South Luangwa National Park (n = 4) and 13, Kafue National Park (n = 2) of Zambia (ZAM) Population, Represent H. a. Capensis Subspecies. The Current Distribution has not been Mapped and the Shading Depicts the Former Range Across Africa [30]
In the study by Okello et al., [30], populations belonging to H.a. kiboko and H.a. capensis subspecies were portrayed as distinct groups radiating from H.a. amphibius, the nominate subspecies. The neighbour-joining phylogenetic tree (Figure 14) however revealed no major geographical partitioning of haplotypes except for a few instances where haplotypes from the same locality clustered together, for example, MFN and MAM haplotypes.
Most of the branches from this tree could not be statistically resolved with more than 50% bootstrap support, due to lack of deep structuring of haplotypes. The statistical parsimony network (Figure 15) showed no clear geographical structuring, except for small star like clusters of haplotypes from Maasai Mara Game Reserve (Kenya), Murchison Falls National Park (Uganda) and Naivasha (Kenya).
Despite lack of clear distinct lineages, most ancestral haplotypes are inferred to be from Murchison Falls National Park (Uganda) population of H.a. amphibius subspecies.
Past Population Expansion of the Species
In recent past expansion, it was noted that the distribution of pair wise nucleotide differences for the total hippopotamus sample, H.a. amphibius and H.a. kiboko subspecies has revealed generally smooth unmoral patterns (Figure 16) characteristic of a population that has undergone a large expansion.
Despite the putative subspecies acknowledged by most taxonomists, the mitochondrial DNA region suggest that hippopotamus has a relatively low variation. This low but significant differentiation among most hippopotamus populations is evidence of a recent population expansion. Overall nucleotide diversity of 1.9% observed in the study by Okello, et al., [30], is relatively low in comparison with diversity indices of other large African mammals from the same region (Table 2).
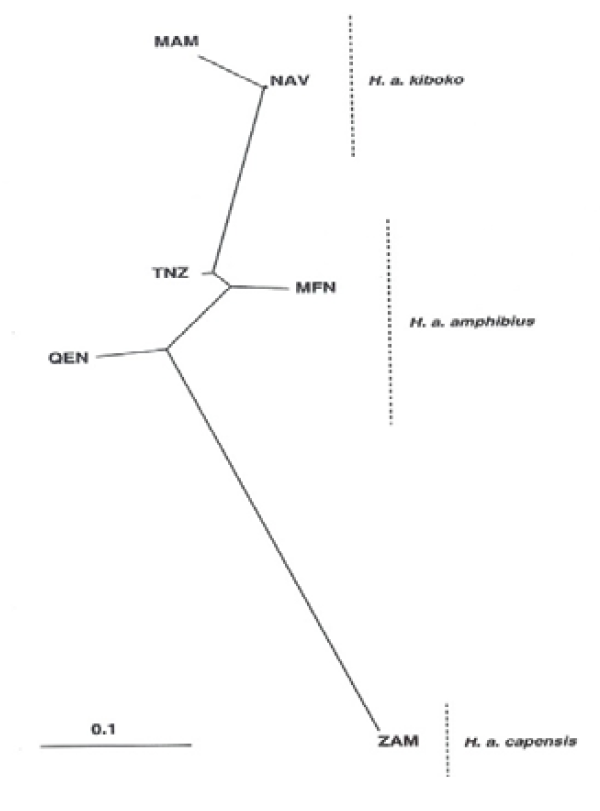
Figure 13: Neighbour-Joining Population Relationships Based on Nei’s net Genetic Distances Among the Six Hippopotamus Populations Sampled [30]
Zambia (ZAM), Queen Elizabeth National Park (QEN), Murchison Falls National Park (MFN), Tanzania (TZN), Naivasha (NAV) and Maasai Mara Game Reserve (MAM)

Figure 14: A Midpoint-Rooted Neighbour – Joining Phylogenetic Tree Based on Maximum -Likelihood Distance Setting and 1, 000 Bootstrap Replicates Among mtDNA Haplotypes of Hippopotamus Amphibius. Zero Branches Were Collapsed Creating Polytomies and Statistical Support for Nodes with ≥50% Boostrap Replicates Are Indicated
Source: [31], Murchison Falls National Park (MNP), Maasai Mara Game Reserve (MAM), Naivasha National Park (NAV), Queen Elizabeth National Park (QEN), Tanzania (TZN), Zambia (ZAM)
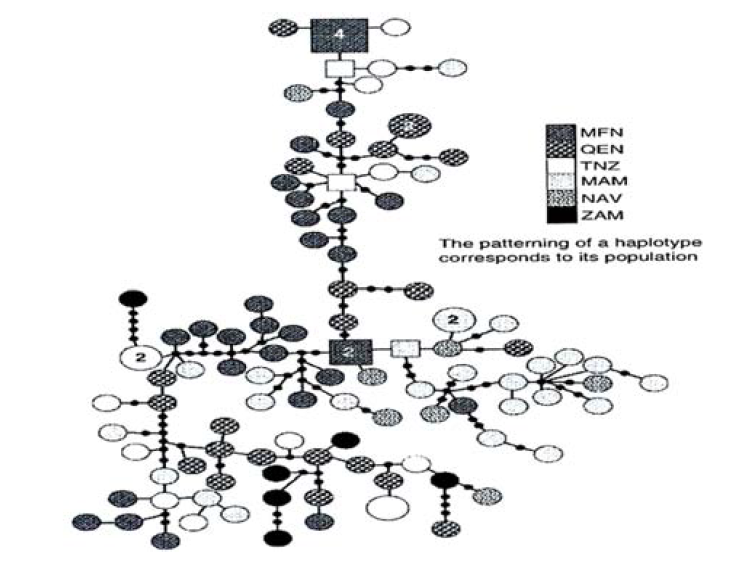
Figure 15: Statistical Parsimony Network Showing Relationships between Hippopotamus mtDNA Haplotypes from Eastern and Southern Africa. Haplotypes Are Represented as Circles and Squares for Ancestral Ones. The Minimum Number of Steps Connecting Parsimoniously Two Haplotypes is Indicated as a Thick Black Square and a Filled Small Circle Represents an Extinct or Missing Haplotype that Might have not been Sampled. The Size of the Square or Circle and Pattern Assigned Correspond to the Haplotype Frequency and Population, Respectively and Numbers in Haplotypes Correspond to the Number of Individual Sharing the Haplotypes [30]
Murchison Falls National Park (MNP), Maasai Mara Game Reserve (MAM), Naivasha National Park (NAV), Queen Elizabeth National Park (QEN), Tanzania (TZN), Zambia (ZAM)
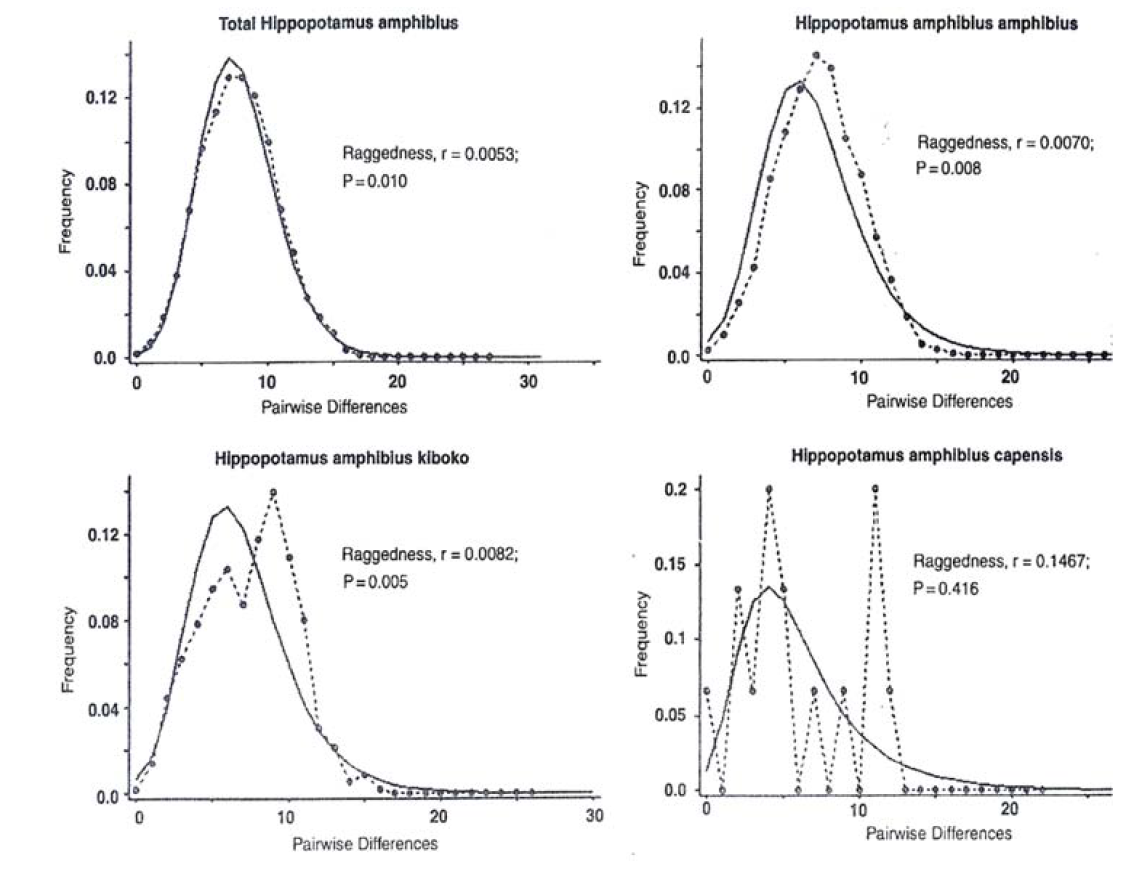
Figure 16: Mismatch Distributions of Mitochondrial DNA Sequences of the Common Hippopotamus based on Pair Wise Nucleotide Differences in the Total Population and Subspecies. Solid Lines in the Curves Indicate the expected Distributions Under Expansion and Dotted Lines Indicate the Observed Distributions Under Population Expansion. The Raggedness Statistics and Corresponding p-values are Given
Source: [30]
Table 2: Comparisons of Diversity Indices of the Hippopotamus with Those of other Large Mammals Studied from the Same Region Based on Nucleotide Sequence of the 5’- end of the Mitochondrial Control Region
| Mammal | Bases | % Var. | (π) | Reference |
| Hippopotamus | 400 | 12.8 | 1.9 | Okello, et al., [30] |
| Warthog | 370 | 7.0 | 4.0 | Muwanika, et al., [31] |
| Elephants | 400 | 8.5 | 2.0 | Nyakaana et al. |
| Impala | 400 | 2.0 | 3.6 | Nersting and Arctander |
| Kobs | 354 | 29.1 | 4.9 | Birungi and Arctander |
Observed nucleotide diversity in hippopotami is only comparable to that in elephants (2.0%); the latter were drastically reduced in numbers in the last few decades due to poaching for ivory [31].
From Table 2 one would expect genetic bottlenecks in hippopotamus in Africa due to low sequence variability and nucleotide diversity observed. This is however, not the case and may be due to relatively short coalescence time since the population expansion event took place and since river systems in Africa are to a large extent interconnected, one would expect to find little or no significant genetic differentiation among these populations. So the resilience of the Luangwa hippo population may not be defined by genetic varions from othger populations but rather environmental factors unique to the Luangwa valley (Figure 16).
Therefore, the variation in population size amomng range states may noit be a consequence of genetic varaiations among populations but rather a factor of the quality of habitat and the effectiveness of the management regimes in those countries. For instance, in areas with protracted civil conflicts, hippo becomes an easy taget because it yields significant quantities of meat and are easy to find since they are confined to water bodies. In Africa for instance, IUCN estimated 157,000 animals in 200, though numbers could be lower now. West Africa had 7,000 split into small groups in over 19 countries. East Africa had about 70,000 with 30,000 in DRC (now less than 20,000), Ethiopia, Sudan and Tanzania, Kenya and Uganda had several thousands. Southern Africa had the largest population size of about 80,000 with Zambia containing the biggest and population (≥40,000) of any country in Africa [13,32]. Other countries in the southern African sub region with large numbers were Mozambique (16,000-20,500), Malawi (10,000), Zimbabwe (6,900) and South Africa (5,000). This number is likely to be higher when surveys are done in Angola. However, owing to loss of grazing land due to cultivation, the numbers are likely to decline in many countries.
Given the potential loss of habitat due to human population expansion and civil strife in many parts of sub-Sahara Africa, the status of hippopotamus can be said to be uncertain in many countries. Civil war in Congo DRC, Liberia and Sierra Leone might have significantly reduced population status. Lewison [32], showed that populations were only increasing in Gabon, Zambia and perhaps Tanzania (Figure 17). From this assessment, it was found fitting that IUCN Species Specialist Group upgraded the species to International Red List of threatened species in 2006. The populations most at risk were the small populations in West Africa, where the distribution continues to be fragmented due to human population expansion and agricultural activities as well as civil wars. The recent attempt by Guinea supported bu other countries to uplist the CITES appendix listing fromn Appendix II to Appendix I on the basis of international trade is not supported by science. The major factors contributing to the loss of hippo in Africa is habiota loss and massive losses due to poaching in areas with civil conflicts and not international trade.
Conducting this review of the hippo population for Zambia was important since it has the largest and growing population than any other range state. Results and recommendations from this review may be helpful for the Zambian party attending the forthcoming conference of the parties to the Convention on International Trade in Endangered Species of WEild Fauna and Flora (CITES) at which a proposal to up lift the Appendix of hippo from Appendix II to Appendix I will be debated.
Current Major Threats
Throughout the range, international trade does not seem to be a major factor that would drive the species to extinction, but rather other factors. For instance, populations of common hippopotamus have declined mainly in response to human disturbances. The species, which was once found through-out sub-Sahara Africa, is now primarily concentrated to Eastern and Southern Africa.
The principal threats to survival are loss of grazing lands mainly due to cultivation and draining of wetland areas, poaching for meat and ivory and wildfire which destroy the habitat [32].
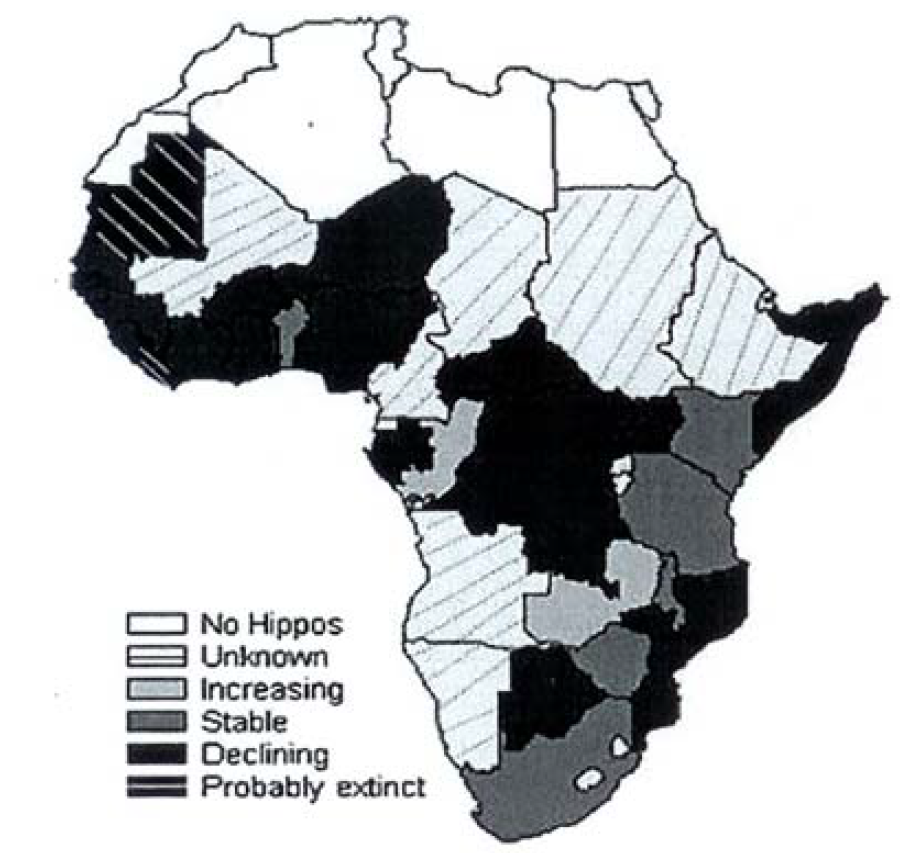
Figure 17: Current Common Hippopotamus Population Status and Trends. Only Congo and Zambia Registered Increasing Populations [32]
Changes might have occurred in the population sizes since then but the patterns are essentaially the same
Poaching for Meat
Adult hippopotamus is hunted for meat to meet the nutritional needs of growing human population [1]. Despite the isolated traditional beliefs in some areas such as the Luangwa valley of Zambia and the Rukwa area in Tanzania where eating hippopotamus meat is believed to cause leprosy, poaching still remains as one of the major causes of population loss in sub-Sahara Africa. Hippopotamus yields a lot of meat, which fetches a lot of money on the black market in urban areas. For instance, in the Democratic Republic of Congo, which was once home to the largest hippopotamus population in Africa, populations have declined by ≥95% in recent years from very intense hunting pressure [32].
Loss of Habitat Due to Agriculture
Information from the IUCN report [33] suggests that in many range states where hippopotami are found, their populations have declined. Such decline has been attributed to anthropogenic activities such as habitat loss as wetlands are converted or impacted by agricultural development [34]. Small-scale habitat loss is most commonly the result of weirs or dams redirecting water from rivers and lakes to crop areas where hippopotamus is not tolerated. In other areas such as large parts of the Kafue system in Zambia wildfires destroy the grassland habitat depleting the food base for the hippopotamus [35]. When anthropogenic activities are considered in conjunction with natural disturbances, even relatively moderate or mild human mediated disturbances may lead to significant population changes [32]. The current expected impacts of global climate change may also affect the species’ population size and distribution in sub-Sahara Africa (personal experience).
International Trade in Hippopotamus Ivory
When one considers the word ‘ivory’ the tusk of the elephant is generally brought into picture. Yet it is just as appropriate to identify the incisors and canines of the hippopotamus as ivory tusks. Elephant and hippopotamus ivory are both primarily composed of dentine and furthermore both represent permanent teeth of continuous growth and hence the designation tusk [36]. In fact, dentine is found in all teeth and is a non-cellular structure with organic and inorganic components. However, it is possible to differentiate between elephant and hippopotamus ivory based on morphological characteristics and structural differences in dentine formation. Some claim for instance, that ivory from forest elephant (Loxodonta africana cyclotis) is harder than that from the bush elephant (Loxodonta africana africana). Similarly, hippopotamus ivory is denser. In the end, though and disregarding enamel and cementum, all ivory ranks between 1.5 and 2.5 on the Mohs scale [37].
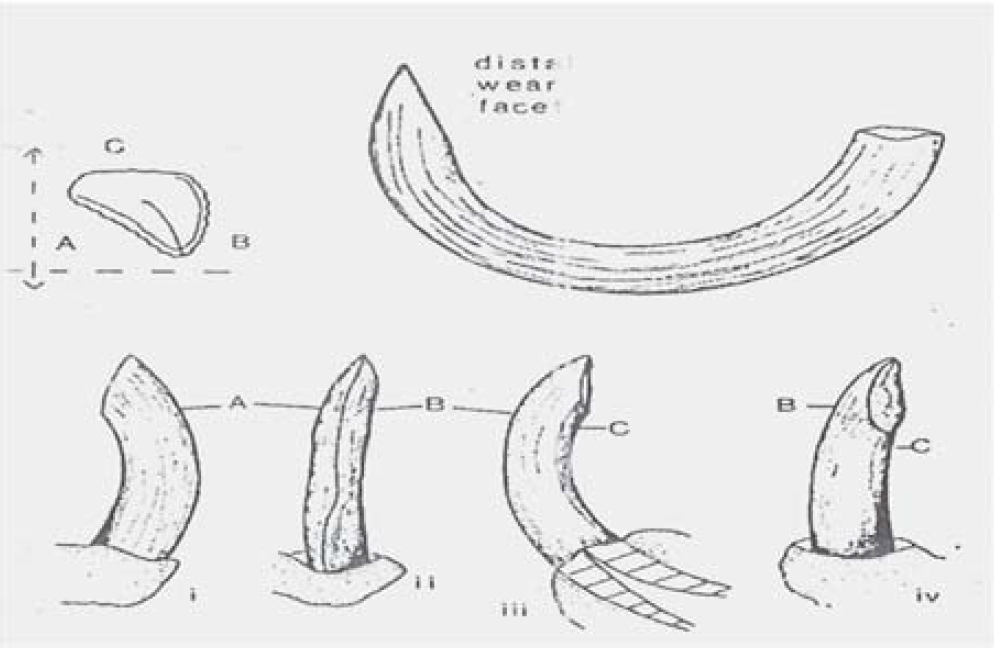
Figure 18a: Characteristics of Hippopotamus Teeth
Source: [37]. A: Represents Outer Surface of the Tooth and ‘C’ Inner Surface of the Tooth
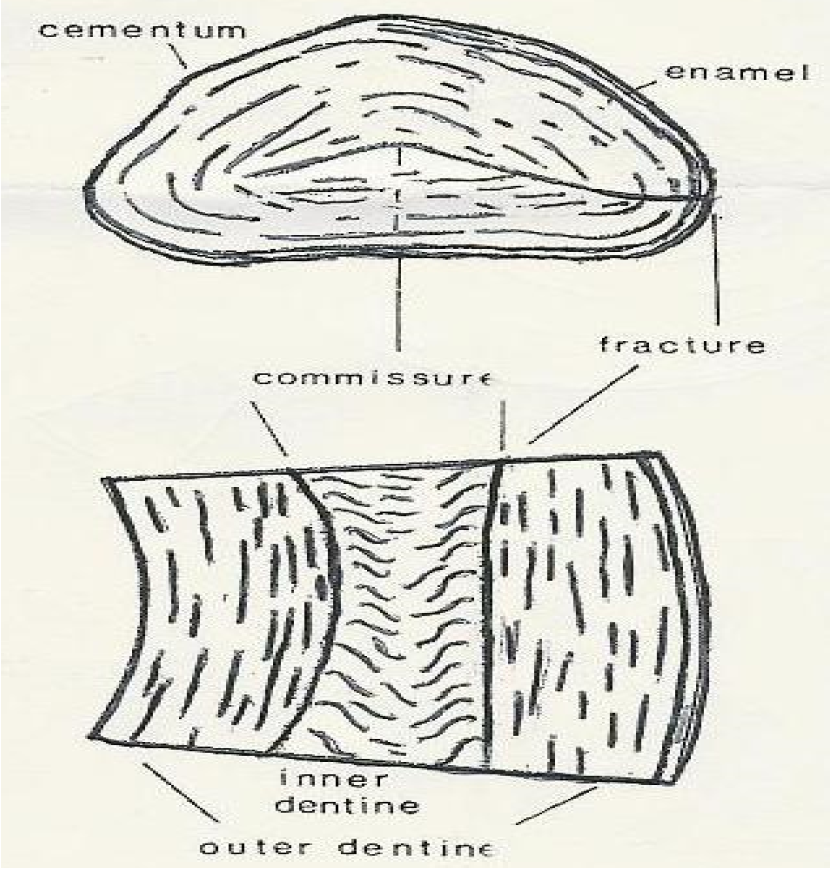
Figure 18b: Lower Canine in Longitudinal Section
Source: Krzyszkowska, [37]
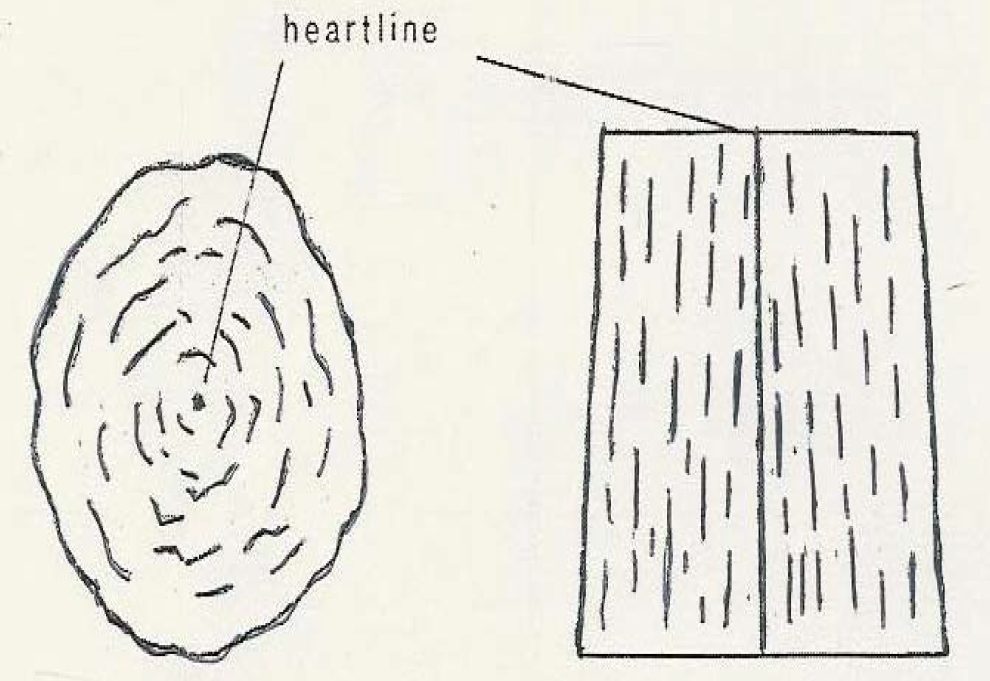
Figure 18c: Hippopotamus Incisor Showing Patterns of Lamellae in Transverse Section (Left and Longitudinal Section (right) [37]
The longitudinal section shows the lamellae running parallel with the surface and curving toward the distal end of the heart line
Historically, hippopotamus ivory has been traded side by side with elephant ivory for centuries, [36] and will likely continue to be traded for as long as the hippopotamus lives. This is because certain attributes inherent in hippopotamus ivory are different from elephant ivory. For instance, hippopotamus ivory is composed of dentine which is formed differently from elephant ivory [13]. Dentine in hippopotamus ivory is recognized as denser and whiter than elephant ivory due to a tighter packing of smaller dential tubules and thus less predisposed to decay [36]. Moreover, where the lamellae in elephant ivory are very regular and even, in the hippopotamus ivory they are wavy and discontinuous [37]. The hippopotamus teeth are also generally smaller than elephant teeth, for instance hippopotamus canines weigh about 2 kg and reach 30 cm in length, with a potential upper limit of 70 cm due to a maximum 40 cm length root. Despite the smaller size however, one hippopotamus skull, yields many pieces by number not by weight of teeth for the ivory market than elephant (Figure 18a, b and c). For this and other reasons, hippopotamus ivory may not in the strict sense be a complete substitute for elephant ivory and vice versa. The whiter color and denser structure could have rendered the hippopotamus ivory more valuable or sought after than elephant ivory in ancient times [36]. If it were not for the nature and shape of hippopotamus tusks which are harder to work with, even today, hippopotamus teeth could have probably been as valuable as in ancient times. For instance, the triangular cross-section and curving nature of the lower canine restricts the designs, which may be fashioned from it [13]. Inlays, discs and lids are ultimately the favoured uses of hippopotamus ivory (Bowman, personal.comm.).
Given this background therefore, international trade in hippopotamus will always persist and may under certain circumstances contribute to the continuing decline in hippopotamus populations in Africa [33]. The global trade in hippopotamus parts includes a wide range of products ranging from skin and leather products to bone, trophies, skulls, feet, shoes and raw carved ivory [33]. An examination conducted by TRAFFIC in 2004 [33], found that the United States of America was a significant importer of these products, with the primary exporting country being Hong Kong [38]. Although this report may now be 18 years old, the pattern may not have changed. Many people claim, including the proponents of the proposal to uplift Appendix listing of hippo from Appendix II to Appendix II of CITES that the contemporary international trade in hippopotamus ivory it differs from the elephant ivory trade in fundamental ways. Most significantly because the hippopotamus is currently only listed in CITES Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) and is otherwise not prohibited from trade by US or international law. This implies that there is an open commercial trade in hippopotamus ivory and other products. That the commercial trade in hippopotamus ivory that existed before the species was listed in CITES Appendix II in 1995 continued afterwards, albeit under closer international monitoring [38]. Williamson [38] also provided statistics on the quantities of hippopotamus ivory imported into the United States of America, between 1995 and 2002. For instance, the USA cleared some 2,479 shipments of hippopotamus ivory in various forms, while refusing to declare 93 others [33]. Although the USA imported far fewer shipments of hippopotamus ivory than elephant ivory during the period 1995-2002, the amount of actual hippopotamus ivory imported far exceeded that of elephant ivory, demonstrating a distinct difference in the pattern of trade between the two commodities [33]. For instance, between 1995 and 2002, the USA reported 6,796 cleared shipments of elephant ivory almost three times the number of cleared hippopotamus ivory shipments. However, the hippopotamus shipments cumulatively represented close to 87,500 individual items as well as 1,436 kg of hippopotamus ivory reported by weight [38]. Hippopotamus ivory tended to be imported in bulk commercial shipments with some hundreds of individual ivory carvings or teeth included in each [33].
Regarding the countries of origin, TRAFFIC, Anon [33] reported that there was by far more information available about the countries of origin for hippopotamus ivory imported in the USA than is known about the origin of elephant ivory. Whereas current legal commercial trade in elephant ivory is largely restricted to antiques for which the original source may no longer be known, the trade in hippopotamus ivory involved a continuous, contemporary stream of ivory from range states to world markets. Tanzania was identified by TRAFFIC, [33] as the lead country of the origin for the vast majority of hippopotamus ivory commercially imported into USA, which is consistent with that nation’s prominent role as an exporter of hippopotamus tusks and teeth. Other nations that featured prominently in the trade included in order of importance, Zimbabwe, South Africa, Uganda, Zambia, Burundi, the Democratic Republic of Congo and Malawi. Of the 810 shipments of carved hippopotamus ivory recorded as entering USA legally between 1995 and 2002, a total of 517 (64%) shipments originated from Tanzania [33,39]. Similarly, Tanzania was recorded as the country of origin for 199 (40%) of the 502 imports which consisted of hippopotamus teeth [33]. Such trends if continued would be a potential threat to the future of the hippopotamus population.
Status of Common Hippopotamus in Zambia
In Zambia, the hippopotamus population benefits from the enhanced protection of the species in a wide network of protected areas covering approximately 31% of the country’s total area of 752,620km2. In the Protected Areas (PAs) major threats to the hippopotamus habitat such as commercial agriculture do not occur. Wildfire is not permitted, although sometimes poachers set fire, or at times fire may start from outside the PA into a GMA or National Park. Generally, protected areas are safe from agricultural activities, which constitute a major threat through habitat loss. The country also has a large number of rivers, lagoons, flood plains and swamps and lakes most of which are located in PAs. This coupled with favorable amount of rainfall ranging from about 600 mm in the south to up to 2,000 mm in the north contribute to forage production and all year round water availability, which support a diverse population of wildlife including hippopotamus.
Plant Species Diversity and hippopotamus Density Distribution
More than twenty-six species of herbs and grasses with more than 5% relative frequency used as food by hippo have been recorded in the short and tall grasslands of the Luangwa alluvial belt [1,22-26]. Preference is skweded towards short grasslands than tall grasslands and often hippopotamus pathways passed through tall grasslands to short grasslands, an indication that short grasslands were utilized more than tall grasslands by hippopotamus (Figure 19). The short body stature and lip morphology of hippo makes it easier for it to feed on short grass lands where it maintains hippo lawns of short grasses leaving tall grasses (Figure). This explains why there was a higher sighting frequency of grazed species in short grasslands than tall grasslands (Figure 6).
In conclusion therefore, we suggest that the abundance and quick recovery of the hippo population in Zambia and particularly the Luangwa Valley (Figure 19) is a blessing in disguise and should not be considered as a curse. A compromise between the two groups; the non-consumptive and consumptive user groups should give eacxh other space as the resource base has the capacity to accommodate both uses. Comments from local and international stake holders regarding how the species should be utilized at national level should be based on science and not popularism or nip mthe bud mentality. Lastly, the proposal to uplift the species from Appendix II to Appnedix I of CITES may not be relavant to Zambia. If parties feel compelled to do so, the best option to be considered perjhaps would be slpilty listing with smeo countries with stable to increasing populaytions mainting it in Appendix II as is the case with the elephant.

Figure 19: Extra Ordinary Large School of Hippo, Luangwa Valley
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. Authors used own funding.
Chomba, C. “Factors regulating the Luangwa (Zambia) hippopotamus population within carrying capacity band.” International Journal of Biodiversity and Conservation, vol. 5, no. 3, 2013, pp. 109–121. https://doi.org/10.5897/IJBC12.023.
McCalough, D.W. Wildlives, Profiles of East African Mammals. African Wildlife Foundation, 1989.
Kingdon, J. The Kingdon Field Guide to African Mammals. A&C Black Publishers, 2008.
Skinner, J.D. and R.H.N. Smithers. The Mammals of the Southern African Subregion. University of Pretoria, 1990.
Saikawa, Y. et al. “Pigment chemistry: The red sweat of the hippopotamus.” Nature, vol. 429, no. 6990, 2004, pp. 363.
Mathee, C.A. et al. “Mining the mammalian genome for artiodactyl systematics.” Systematic Biology, vol. 50, 2001, pp. 367–390.
Amrine-Madisen, H. et al. “A new phylogenetic marker, apolipoprotein B, provides compelling evidence for eutherian relationships.” Phylogenetic Evolution, vol. 28, 2003, pp. 225–240.
Haltenorth, T. and H. Diller. A Field Guide to the Mammals of Africa Including Madagascar. Collins, 1988.
Dorst, J. and P. Dandelot. A Field Guide to the Larger Mammals of Africa. Collins, 1970.
Corbet, G.B. “The taxonomic status of the pygmy hippopotamus, Cheropsis liberiensis from the Niger Delta.” Journal of Zoology, vol. 158, 1969, pp. 387–394.
Bosman, P. and A. Hall-Martin. Elephants of Africa. New Holland, 1989.
Boisserie, J.R. “The phylogeny and taxonomy of hippopotamidae (Mammalia: Artiodactyla): A review based on morphology and cladistic analysis.” Zoology, vol. 143, 2005, pp. 1–26.
Eltringham, S.K. The Hippos. Academic Press, 1999.
Stuart, C. and T. Stuart. Field Guide to Mammals of Southern Africa. Struik Publishers, 2001.
Laws, R.M. et al. “Observations on reproduction in the hippopotamus Hippopotamus amphibius.” Symposium on Zoology, vol. 15, 1966, pp. 117–140.
Sayer, J.A. and A.M. Rhaka. “The age of puberty in the hippopotamus (Hippopotamus amphibius) in the Luangwa river in eastern Zambia.” East African Wildlife Journal, vol. 12, 1974, pp. 227–232.
Chomba, C. et al. “Does shore length determine population size and density distribution of hippo in Zambia?” Journal of Ecology and the Natural Environment, vol. 4, no. 6, 2013, pp. 150–153.
Chomba, C. et al. “Has the Luangwa (Zambia) hippopotamus (Hippopotamus amphibius) population stabilized after reaching its asymptote? What are the management implications?” Open Journal of Ecology, vol. 3, no. 6, 2013, pp. 395–406.
Chomba, C. et al. “Does the Luangwa hippo population size and density distribution vary between upper and lower study blocks? What are the management implications?” Open Journal of Ecology, vol. 4, no. 5, 2013, pp. 262–280. http://www.scirp.org/journal/OJE/.
Asdell, S.A. Patterns of Mammalian Reproduction. Collins, 1964.
Marshall, P.J. and J.A. Sayer. "Population ecology and response of a hippopotamus population in eastern Zambia." The Journal of Applied Ecology, vol. 13, 1976, pp. 391–403.
Chomba, C. and H. Chabwela. "Does mortality impact negatively on the hippopotamus population size and density in Luangwa river, Zambia? A historic perspective." Global Journal of Biology, Agriculture and Health Sciences, vol. 5, no. 4, 2016, pp. 7–15. www.gifre.org.
Chomba, C. "Does kidney fat index determine body condition of the common hippopotamus (Hippopotamus amphibius) in Luangwa river Zambia?" Global Journal of Biology, Agriculture and Health Sciences, vol. 5, no. 4, 2016, pp. 16–23. www.gifre.org.
Chomba, C. "Population structure of the common hippopotamus (Hippopotamus amphibius) in Luangwa river Zambia." Global Journal of Biology, Agriculture and Health Sciences, vol. 5, no. 4, 2016, pp. 29–39. www.gifre.org.
Chomba, C. "Does kidney fat index determine body condition of the common hippopotamus (Hippopotamus amphibius) in Luangwa river Zambia?" Global Journal of Biology, Agriculture and Health Sciences, vol. 5, no. 4, 2016, pp. 16–23. www.gifre.org.
Chomba, C. "Population structure of the common hippopotamus (Hippopotamus amphibius) in Luangwa river Zambia." Global Journal of Biology, Agriculture and Health Sciences, vol. 5, no. 4, 2016, pp. 29–39. www.gifre.org.
Plowright, W. et al. "Serological evidence for the susceptibility of the hippopotamus (Hippopotamus amphibius L) to natural infection with rinderpest virus." Journal of Hygiene, vol. 62, 1964, pp. 329–336.
Tembo, A. "Population status of the hippopotamus on the Luangwa river, Zambia." African Journal of Ecology, vol. 25, 1987, pp. 71–77.
Stocher, J.S.B. et al. "The diet of the hippopotamus in ndumu game reserve, natal, as determined by Faecal analysis." South African Journal of Wildlife Resources, vol. 8, 1978, pp. 1–11.
Okello, J.B.A. et al. "Mitochondrial DNA variation of the common hippopotamus: evidence for a recent population expansion." Heredity, vol. 95, 2005, pp. 206–215.
Muwanika, V.B. et al. "Phylogeography and population structure of the common warthog (Phacochoerus africanus) inferred from variation in mitochondrial DNA sequences and microsatellite loci." Heredity, vol. 91, 2003, pp. 361–372.
Lewison, R. "Population responses to natural and human–mediated disturbances: Assessing the vulnerability of the common hippopotamus (Hippopotamus amphibius)." African Journal of Ecology, vol. 45, 2007, pp. 407–415.
Anon. Report to Zambia Wildlife Authority on the Hippopotamus Survey. Chinzombo Research Centre, 2004.
Smuts, G.L. and I.J. Whyte. "Relationships between reproduction and environment in the hippopotamus in the Kruger national park." Koedoe, vol. 24, 1981, pp. 169–185.
Anon. South Luangwa Area Management Annual Report. Zambia Wildlife Authority, 2007.
Lafrenz, A.K. "Tracing the source of the elephant and hippopotamus ivory from the 14th century B.C. uluburu shipwreck: The archeological, historical and isotopical evidence." MA thesis, University of Florida, 2003.
Krzyszkowska, O.H. Ivory and Related Materials: An Illustrated Guide. Classical handbook 3, Institute of Classical Studies, 1990.
Williamson, B.R. "The condition and nutrition of elephants in Wankie national park." Arnoldia Rhodesia, vol. 7, no. 12, 1975, pp. 1–20.
James, E. "The influence of elephant ivory trade ban on population of common hippo (Hippopotamus amphibius): A case study of Katavi-Rukwa-Lukwati ecosystem, Tanzania." MSc thesis, University of Dar es Salaam, 2007.
Anon. Report to Zambia Wildlife Authority on the Hippopotamus Survey. Chinzombo Research Centre, 2008.
Sichingabula, H. "Hydrology, character of channel changes on luangwa river, their impacts and implications on tourism, south Luangwa national park and adjacent game management areas." Report prepared for EDF/NPWS towards the development of a General Management Plan for South Luangwa National Park and Adjacent Game Management Areas, 1998.
Chomba, C. et al. "The population status of hippo along the 165km stretch of the Luangwa river, Zambia." Journal of Ecology and the Natural Environment, vol. 4, no. 9, 2011, pp. 247–257.
Ansell, W.F.H. "Hippo census on the Luangwa river." Puku, vol. 3, 1965, pp. 15–27.
Arman, P. and C.R. Field. "Digestion in the hippopotamus." East African Wildlife Journal, vol. 11, 1973, pp. 9–17.
Arctander, P. et al. "Phylogeography of three closely related African Bovids (tribe alcelaphini)." Molecular Biology of Evolution, vol. 16, 1999, pp. 1724–1739.
Attwell, R.I.G. "Surveying Luangwa Hippo." Puku, vol. 1, 1963, pp. 29–49.
Bronson, F.H. and B.E. Eleftherriou. "Adrenal response to fighting in mice: Separation of physical and physiological causes." Science, vol. 147, 1965, pp. 627–628.
Coryndon, S.C. "The extent of variation in fossil hippopotamus from Africa." Symposium on Zoology, vol. 26, 1970, pp. 135–147.
Coryndon, S.C. "Fossil hippopotamidae from pliocene/pleistocene successions of the rudolph basin." Paper for Conference on Stratigraphy, Paleontology and Evolution, Wenner Grenn Foundation, 1973.
Coryndon, S.C. "The taxonomy and nomenclature of the hippopotamidae (Mammalia, Artiodactyla) and a description of two fossil species." Proc. K. Nederlandse Akad. Wetnsch. Series. B, vol. 80, 1977, pp. 61–88.
Grubb, P. "The afrotropical hippopotamus and Hexaprotodon: Taxonomy and description." Pigs, Peccaries and Hippos, edited by W.R.L. Oliver, IUCN, 1993.
Harpending, H.C. "Signature of ancient population growth in a low–resolution mitochondrial DNA mismatch distribution." Human Biology, vol. 66, 1994, pp. 591–600.
Klingel, H. "The social organization and behaviour of the Hippopotamus amphibius." African Wildlife Research and Management, edited by F.I.B. Kayanja and E.L. Edroma, International Council of Scientific Unions, 1991.
Laws, R.M. and G. Clough. "Observations on reproduction in hippopotamus (Hippopotamus amphibius L.)." Comparative Biology of Reproduction in Mammals, vol. 117, 1965, pp. 40.
Laws, R.M. "Dentition and ageing of the hippopotamus." East African Wildlife Journal, vol. 6, 1968, pp. 19–52.
Olivier, R.C.D. and W.A. Laurie. "Habitat utilization of hippopotamus in the Mara river." East African Wildlife Journal, vol. 12, 1974, pp. 249–272.
Wright, P.G. "Thermoregulation in the hippopotamus on land." South African Journal of Zoology, vol. 22, 1987, pp. 237–242.
